Important explanation on updated age of site: The actual age of the El Pueblo ichnotelmafacies (swamp facies) has been updated to late Carboniferous or Pennsylvanian age. A Kasimovian stage age (305 mya) seems to be emerging based on the high diversity and high population of tetrapods in Kasimovian forested swamps (as parental to the El Pueblo lowland swamp). The Kasimovian stage was an early stage evolution age that might explain a coexistence of small ichno-Dimetrodons and very large ichno amphibians in the El Pueblo Swamp. An early Permian age no longer applies. Fossils found many years after its discovery revealed that the El Pueblo swamp existed at a time when the upland was still low and giant dragonflies were still flying around. Fauna of the EL Pueblo fauna did evolve further and transferred unto the early permian at the age level of the base of Glorieta Mesa as US geologists estimated. I was of a similar opinion that the Glorieta-Mesa stratifications contain the entire time-scale of the Permian Period. I have uploaded a Kasimovian-stage abstract to academiaedu.com (Henry Ortiz). Every effort was made to conform the age of the El Pueblo ichnotelmafacies to Wolfcampian early Permian as designated by state workers in 1990. Revisions in geological and paleontological abstracts are common because of changing paleo-evidence, many unknowns, as well as biases and variable interpretations. I will continue a late-Pennsylvanian-age edit of this blog as time allows. Please excuse the inconvenience. Henry Ortiz 6/15/2024.
By Henry Ortiz, former flagstone-quarry worker
 |
Four trackways in one, largest footprint
fits inside a dime. |
The El Pueblo swamp held a diverse group of lizard-like animals called tetrapods ranging from hatchlings to adult size. The photo above shows a slab containing small trackways. This swamp, like any other wild prey-predator environment, attracted such hunters as Dimetrodon and a host of other large tetrapods capable of ranging from swamp to swamp.
😞Trying to Mitigate the Confusion😊
This blog has been well received by many fossil enthusiasts as well as by researchers. People are forgiving when they spot mistakes or confusing descriptions. I am in the process of editing this blog for easier understanding from the reader's perspective. First of all I had been referring to the El Pueblo early permian swamp facies as EPEPSF, in order to save words as the saying goes. That will be changed to either El Pueblo ichnotelmafacies or the El Pueblo facies or the El Pueblo swamp. Ichnotelmafacies or anything with facies refers to the present rocky remnants of the swamp, whereas the El Pueblo swamp refers to the bygone vital and functioning swamp of over 300 million years ago. Also the second biological name for each plant or footprint fossil will be elpueblus, referring to locality and not necessarily to a new species. This method was used by Tilton in1931 in order to distinguish Dimetrodon berea (Berea, West Virginia) from other Dimetrodon footprints. Source of fossils is important in order to establish all the different source localities. Tilton's method worked and now ichno-Dimetrodon berea elpueblus confirms two localities. Also the prefix ichno will be used occasionally as an affirmation that no skeletal examples exist. The entire El Pueblo facies is an ichno (trace path) facies with a high ferrous content. The El Pueblo swamp did not contain enough calcium carbonate to neutralize oxidation long enough to prevent decay of skeletal remains and of most plants.
The El Pueblo Early Permian Swamp Facies of
Northeastern New Mexico
The El Pueblo swamp facies has elucidated the true ichnotaxa that transferred onto the land during the Pennsylvanian Epoch. It presents a recently discovered world-class example of the roots that launched terrestrial evolution. It is also an example of Pangean plate tectonic forces that apparently expedited a transfer of inland marine flora and fauna into terrestrial lakes and swamps. Although there is evidence that the El Pueblo swamp began at the Pennsylvanian juncture its age description will continue as Artinskian Early Permian so as not to confuse readers. The El Pueblo ichnotelmafacies (swamp facies) exhibits fossils of the late Carboniferous age (305 MYA). An abstract entitled When the Geologic Time Scale Does not Match the Evidence was uploaded to Academia.edu. A distinction from Mississippian (sea swamps and ponds) to Pennsylvanian (terrestrial) is at the root of this distinction. Both end-Pennsylvanian (the preceding Epoch) or very early Permian, remain viable time designations for the El Pueblo Ichnotelmafacies (swamp facies).
A World of Ferns
Ferns were the main food source for herbivore animals of the late Carboniferous and early Permian. Because ferns are soft plants that do not resist decay, a lack of fern impressions was confusing at the time the El Pueblo ichnotelmafacies (footprints-swamp-rockfaces) was discovered. Thanks to the subtraction of ferrous tint via the oxygen in ferns, many stands of large ferns were discovered via oxidative fossils along the main flowway that provided runoff into the El Pueblo swamp. This proved that the swamp perimeters and flowways were a lush place where many animals fed on ferns as one of the root tiers of the food chain. The basic food tier was plankton. The El Pueblo swamp finally fit the mold of previously discovered fern and conifer environments of the early Permian Period. In the El Pueblo swamp trajectory (305 MYA) the early Permian Period was simply extended biologically for about five million years because of expedited burial of the inland sea. After five million years (which is a rocks throw in geological time) early Permian evolution continued. The whole thing amounts to a terrestrial head start for swamp animals on the lowland. After the El Pueblo swamp became buried in sediments, both flora and fauna progressed down the alluvial valley within the advancing Permian Period. A Triassic dinosaur was found just twenty miles downriver at a place where the upland plain and the Pecos river are separated by a low difference in altitude. During the early Permian Period the Pecos River did not exist. Although there was runoff from mountains the lowland was not yet inclined toward the south as in the present. Progressive subsidence at the base of mountains was holding back water in the form of lakes and swamps. Eventually a trough fault in a present north to south direction (the future valley) provided the drainage that established a string of low-lying swamps such as the El Pueblo swamp (Henry Ortiz, 2023)
The Sangre de Cristo Orogeny
Click on image to enlarge
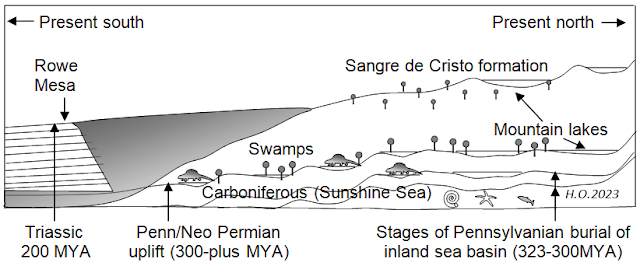
Above diagram: The Pecos Valley trajectory of evolution was from north to south. The transitional orogeny from marine to terrestrial is sometimes referred to as a montane type as opposed to a coastal type. (In coastal transitions some animals that lived in swampy areas close to the sea actually reverted back to a marine life.) Uplift episodes in the upper Pecos Valley were many, and usually very slow. Sometimes uplifts were sudden due to movement of the igneous basement and volcanic upthrust. Sudden uplift resulted in increased gradient and a massive downslide of sediments and rocks. The conveyance of sediments and rocks from the upland to the lowland (normally) were by rainfall and flooding. After the separation of continents and the end of Pangea, surficial deformation of the earth crust abated almost entirely. Once at 1000 feet above sea level the El Pueblo ichnotelmafacies is now located at 6200 feet above sea level. The great erosive phase, which divided the mesaland, began after the beginning of the Triassic Period. This juncture marks the final tectonic uplift that tilted the terrains toward the present south; the separation of continents; and the end of mesa-plain building. Shortly thereafter the Pecos River and tributaries became the incessant conveyor of sediments and rocks, eventually gouging a deep valley that is now the upper Pecos Valley.
Background on the
El Pueblo Ichnotelmafacies
The El Pueblo ichnotelmafacies of northeastern New Mexico, US, is located in the Upper Pecos Valley near north-central New Mexico. El Pueblo refers to "the pueblo", a nearby pueblo ruin of the ancient Anasazi. I discovered the fossil site in 1984 while prospecting for commercial flagstone. The facies is composed of weathered and fluvially transferred granitic sediments from the Sangre de Cristo ancestral (Ouachita-Marathon orogen) formation. The ancestral orogeny was the result of collision between the South American and African continents with the North American Continent. Continental-crust subduction ensued, thickening the local earth crust, and creating repetitive granitic uplifts that weathered into clayey sediments. Uplifts caused by continental collisions displaced the inland sea, creating alluvial aprons and a periphery of swamps. The inland sea parental to the El Pueblo early Permian Swamp, which I named the San Ysidro Inland sea because of marine exposures in namesake villages, was literally emptied out by continuous uplifting. (There is some evidence that the parental sea was called the Sunshine Sea by past geologists and I wish that name would persevere.)
Only trace fossils have been found in the El Pueblo facies so far. The general rule is that bones are not found where footprints are found and vice versa. Some fossils, such as large dragonfly footprints and large three and four digit amphibian footprints, hint of a Pennsylvanian age. These fossils correspond to volcanic episodes and immense uplift during the end Carboniferous that emptied sea lakes and expedited a transfer of fauna into the lowland. Deep-well oil exploration revealed a relatively shallow Pennsylvanian sea and very marginal carbon content. Furthermore, rampant volcanic venting cooked what little oil was present, if any, (e.g. Cone Plant lake which was incinerated by volcanic venting [Henry Ortiz 2013]).
The Terrestrial Environment of
the Early Permian
The main terrestrial animals of the Pennsylvanian and early Permian were lizard-like animals called tetrapods. The El Pueblo swampsite facies has produced a large number of tetrapod footprints and plant impressions now believed to be five million years earlier than the early Permian. Recent research has finally offered an explanation for the rich variety of fauna and flora that was present in the El Pueblo early Permian swamp. Not only were the proper geological factors conducive to the recording of the many impressions, but the site was located relatively close to the western seacoast and almost precisely at the equator. Furthermore, a shallow remnant-like inland sea persevered south of the ancestral uplifts. Rainfall was localized and repetitive. Monsoons from the main ocean replenished the outlying marine basins. The proper climatic and geological conditions for the evolution of terrestrial fauna was in place. Vegetation was in plentiful supply, providing a high rate of CO2 conversion into oxygen. There is evidence that tetrapodic wildlife actually became smaller at the Carboniferous-Permian boundary, which in essence was a change from the inland marine to the terrestrial swamp environment. By the early Permian Period tetrapods of the El Pueblo swamp-trajectory had become quite big, many much bigger than during the Carboniferous Period.
Click on images to enlarge
Use standard word-find functions on your PC
A Segment of the Upper Pecos
Late Pennsylvanian to Trans-Permian
Evolution Pathway

Above Google-map image shows the upper Pecos Valley trajectory at about thirty miles from the Sangre de Cristo mountain range to near Sena, New Mexico. Although the entire early Permian alluvial plane and paleo-swamp trajectory (now the upper Pecos River path) was of a redbed composition the peaks at the bottom of the photo now appear to be white in color. That is because these mesa peaks are post-Permian. The concreted erosion-protective caps consist of quartzite lake-bottom sediments now broken into light-colored talus that surround the peaks. After the early Permian trajectory became buried, quartzitic sediments filled the lake trajectories hundreds of feet above the El Pueblo early Permian swampfacies. Because pink biotite granite is usually a soft shallow granite the change from pink-granite sediments to quartzitic sands means deeper-granite uplift during the end of the early Permian and during the Triassic Period. The supply of pink-biotite granite, although in colossal supply from continual subduction phases and sea-bottom accumulations (a containment geologically referred to as the Taos trough) had diminished. The change in sediments also marked a new uplift phase whereby it is possible that light-colored sand accumulations were dislodged from upland quartzitic escarpments. Along with a decrease in redbed alluvials, long-term-underwater hematite expenditure and silication also contributed to light-colored tops, both at the source and throughout the detrital or clastic process. The most important factor in sedimentary pigmentation is the fact that iron gravitated along the alluvial clay valley at different intervals, and remains the main determinant of evolution trajectories. Evolution environments at the limonite-quartzitic level are limited.
Changes from redbeds to quartzitic lake-bottoms seem to indicate a biological impoverishment with a survival avenue for both flora and fauna more prominent along the relatively narrow confines of the redbed alluvial valley coursing south away from the upland. This process, at least in part, demonstrates the natural geological and biological adjustments that were occurring, and that partially explain a Permian extinction or adjustment. The top of mesas contain weathered quartzite particles mixed in with another deposition of red beds, again from weathered reddish granite on the upland, assuring a last deposition of fertile medium for the future growth of evergreens and grasses. A presence of river rock atop mesas proves terrestrial connectivity with the mountainous upland during end-Permian and Triassic times. Apparently the alluvial-clay valley rose along with the flanking mesaland until at long last, dome uplifting created sufficient gradient for the upland streams to further their course southward, repeatedly cutting downward during heavy rains.
The bio-geological column in the area as recorded in deep-well cores shows that the late Carboniferous inland sea (with overlying Permian strata at many points) extended hundreds of miles to the south. It is likely that ancestral faults determined the target areas of extreme uplifts as well as the path of the local alluvial valleys. The El Pueblo early Permian swampfacies is located to the left of the word Pueblo (in small letters, click image to enlarge). The evolution trajectory shown ranges from Late Carboniferous at mountain base to neo-Triassic at top of light-colored peaks. The evolution column was transitional along the alluvial valley trajectory (which is why the term trajectory was adapted). As the process of aggradation and swamp burial progressed southward, swamp taxa was surviving by relocating further south (e.g.,Triassic Dinosaur [Alex Velasquez] 20 miles downriver from the El Pueblo swamp). It is theorized that the final uplift phase occurred roughly 100 million years ago, finalizing the higher horizon of the Sangre de Cristo mountains and ushering in the great erosive phase that at the present has uncovered the early Permian trajectory. Without tectonic pressure from Pangea I have my doubts about this theory. Furthermore, river rock atop Rowe Mesa points to connectivity with the mountains and and a great potential for a long term erosive phase. While erosion has increased, the weathering rate of granite decreased considerably as the continent drifted north of the wet-tropical equatorial latitudes of the Paleozoic. There is debate as to whether or not the southern rocky mountains are still rising. Hard to tell if they are. If they are, ancestral faulting, slight plutonic thrust and increasing summer temperatures would be at the center of causation. Ring-of-fire volcanic activity, which might be exerting lateral pressure on continents, cannot be dismissed completely as a cause of slight growth of mountain ranges. Slight tremors have been felt in the area for a long time. The thirty-mile thick crust in the area undoubtedly hides many geological secrets of long, long ago. Yet, massive uplift, aggregation, and erosion can primarily be attributed to plate-tectonic forces of Pangea, the supercontinent (Henry Ortiz, 2019).
Figuring Out Early Permian Footprints

Above photo shows footprints tentatively identified as those of a snake. If it weren't for the work of past paleontologists, we'd be lost in our attempts to figure out footprints, at least to a credible degree. Even the names they applied to footprints that lack skeletal reference are a good "handle" by which to identify footprints. In the example above a vague conclusion can be drawn as to this set of footprints on the fact that, according to past paleontologists, several types of snakes existed during the early Permian. This fossil was found at the alluvial-clay valley, the lowest point of the El Pueblo swamp trajectory. Not a bad place for snakes, which in the early Permian were essentially water snakes. But some of these snakes had four feet (Lepospondyls). The intrinsic long narrow design was of course for access between close roots and dense vegetation. But then there were also long salamander-type tetrapods that could flex their bodies, creating footprints that were facing each other. These footprints are quite big and very close together, possibly from limbs attached to a narrow trunk. Bottom line: These footprints need some more figuring out.
Newly Discovered Footprint from the
El Pueblo Early Permian Facies.
How about that?! Three digits, five claws. The long pointed claw in the center of the above footprint surely had some kind of utility, either for pick-digging or for stabbing, or both. If there was an early Permian earth-worm digger, this trackmaker was probably it. The El Pueblo facies continues to produce interesting ichnological evidence that rivals or surpasses footprint collections in museums and scientific journals (Henry Ortiz, 1/2021).
Mineralogical Composition of Trace Fossils
From the El Pueblo Swamp Facies
Trace fossils from the El Pueblo ichnoswampfacies are composed of an arkose. An arkose rock (or ground content) is often referred to as having been derived from granite. In the El Pueblo facies the mineralogical content closely reflects the makeup of pink granite because of pigmentation from both a detrital ferrous influence and from pink feldspar. The pink feldspar also has an influence of ferrous oxides. Arkose consists of quartz as the main mineral with a lesser amount of feldspar and mica. In the El Pueblo ichnoswampfacies the ferrous content exists as a pigmentation in an ionic bonding (some say metallic bonding) with the other minerals, usually delivered in a suspension of clayey wash. That ferrous content derives from volcanic venting that occurred during plate collisions and upthrust of mountain ranges. As important as the oxygen delivery agent in the bloodstream, a ferrous content as hematite merged perfectly with the arkose, ensuring aqueous oxygen appropriation in lakes, flowways and swamps. One of the first discoveries in the El Pueblo facies was that off-white fossils at some points of the swamp underwent a subtraction of pink pigmentation through various subtractive-oxidative processes. Mineralogical bonding, either igneous or not, involves attraction and reception (and often slow or abrupt unbonding, as in weathering, decay, or natural disintegration). The union and disunion of minerals is the stuff of life and a component of the God-particle theory. The reason that the layering of the El Pueblo facies is mainly a pink arkose is because early swamps were precisely formed from weathering pink granite, the exact proper medium for the evolution of redbed swamps. Likewise, inland marine environments depended heavily on an arkose content. A colossal amount of granite was brought up by uplifts as life began to change from the inland marine to the terrestrial. (Henry Ortiz, 2020).
 |
| El Puerto means the Pass |
View from the south from atop El Puerto Mesa which is a continuum of Rowe Mesa. The El Pueblo early Permian swamp facies is located at approximately right-center of photo. Rowe Mesa is on upper left. The base of the nearest mountains is composed of end-Pennsylvanian limestone strata from the local inland sea. Sedimentary mass is mostly from eroded and weathered granite that began to uplift since the end of the Carboniferous Period within a tropical climate. During the early Permian the mesas did not exist. The vast volume of granitic sediments that weathered on the upland and that were transferred toward the south for scores of miles through fluvial action negates the popular theory of long-term droughts. Although the law of averages and incidental occurrence would allow for an occasional drought, the overall evidence from the El Pueblo geological trajectory points to repeated and voluminous rainfall that transferred the same sediments that now form thick and dense formations that cover hundreds of square miles. These formations, interspersed by thick layers of alluvial mass wasting, are visible on the sides of the local foothills and mesas.
Life Before The El Pueblo Early
Permian Swamp
Above photo shows fossils from Cone-Plant Lake. Because of uplift and volcanism life could be rough at the end of the Carboniferous Period. The three tetrapod mandibles shown above are remains from abrupt mortality that occurred in an artesian lake before the El Pueblo early Permian swamp existed. These three tetrapods were among the first arrivals north of the El Pueblo early Permian swamp. As the upland uplifted many lakes, ponds and swamps were emptied out or filled up with sediments. Many swamp lakes that survived were overpopulated. This destruction, along with higher gradients, was forcing tetrapodic life into troughs along the base of the upland. Unfortunately the artesian lakes that formed at the base of the upland were dangerous places. Cone plant lake, dotted with early conifer, calamite (horsetails) and cordaites trees, existed at the Pennsylvanian-Permian boundary of about 310 million years ago. Because there are no signs of marine invertebrates in the remnants of Cone Plant Lake, the biological age suggests an initial or expedited neo-Permian terrestrial-life stage. Gases from volcanic venting found their way into the artesian conduits (deep crevices and strata leading from upland lakes), incinerating the entire lake. Trees exploded, and roots burned below the mud. The water from the lake probably evaporated into a cloud of super-heated steam. Tetrapods and other animals in the lake perished. The high presence of finely masticated bones embedded in the sparse limestone can be attributed to all of the dentitions shown above, although the middle mandible (short thick skull) might be from a heavy catcher-biter such as Dimetrodon. The inferential evidence suggests that the mastication of invertebrates during marine life out of necessity transferred to the mastication of bony material at the neo-Permian. The long-jawed "crocodilian catcher entity" is suggested in the lower mandible. The top mandible suggests a strong thick skull of another strong biter like Dimetrodon. Not too much evidence of a herbivore here, although occasional omnivorism was possible during the late Pennsylvanian. In my opinion the three dentitions suggest an entity of tear or crush and then swallow, much like the canine or feline animals of the wild. The absence of large bones suggest that these mandibles belonged to smaller tetrapods that were flood-transferred along a principal flowway from an upland lake to the bottomland. These prodigy reflects successful survival and highly enumerating hunter-tetrapods that were at the forefront of terrestrial evolution. They probably presaged some of the larger reptilian ichnology in the El Pueblo facies. Unfortunately, many bottomland as well as upland lakes were deadly places during the volcanic onslaught of the end-Pennsylvanian Period. The mandibles might be of infantile age. Adult size of many tetrapods during the Pennsylvanian-Permian boundary Period remains uncertain.
The "Cone Plant" From Cone Plant Lake
Above photo shows a cone-plant-like fossil which turned out to be the root portion of a calamite tree. (When I first saw this fossil I thought I had found the cone plant of mythical lore.) It is lacking several layers. This is part of a young calamite tree that was charred and toppled by volcanic venting. With the top portion of the tree incinerated this bottom portion sank into the mud where it fossilized into stone. Cone Plant lake was highly rhizomatous. The calamite trees vastly outnumbered the Walchia conifers and cordaites trees.
An Overlap in Geological Time

Above photo shows the traces of two ancient sea lilies. The one on the left is from the El Pueblo early Permian swampfacies. The one on the right is from the parental Pennsylvanian upland. Both were detrital and lacking a stem when their traces were recorded. We tend to forget that biological and geological Periods (biostratigraphy) sometimes overlapped because evolution did not occur in one sudden change. The trace on the left is from the head of a sea lily that was flood-imported from the upland along a flowway into the El Pueblo swamp. This happened at a time when the vestiges of marine lakes still existed on the upland. The upland was much lower than it is today, although colossal amounts of weathered sediments had already been deposited southward. A long-term cycle of uplift and weathering was in effect. These two sea lilies conjure an image of swamps leading down from marine lakes that were becoming more and more shallow as uplift from intercontinental collision continued. Uplifting continued even after the continents separated because tectonic and subductive volcanism, once triggered, tends to recur. Some have called these events a symphony of time but the destruction of inland-sea life was incalculable. Countless types of invertebrates perished. Sea lilies and many other marine plants did not survive in the ever-aggrading swamps and flowways of the early Permian. But the natural mandate was that the inland would diversify in elevations. From mountains to prairies to main oceans. However, nature's hand assured the survival of future biological environments. Thrust faulting of the Permian and Triassic Periods insured the survival of stepped gradients. After sea-lake vestiges disappeared rain-dependent lakes persisted on the upland, some which are now favorite fishing spots accessible only by long hikes through rugged terrain. (Updated 7/22/2020, Henry Ortiz)
Did large plants and trees of the El Pueblo ichnotelmafacies have annular rings?
For a long time we have inferred that all tree rings are annular rings. The climate of the El Pueblo swamp was tropical. Plants grew year-round because of very small variations in rainfall or temperature. The above photo shows a fossilized partial stem of a large plant from the El Pueblo ichnotelmafacies (swamp facies), possibly a cycad. The rings can be better described as growth rings. This type of growth is designed to store water in the pulp separating each growth ring. It remains uncertain if each tree ring grew in accordance to a natural yearly cycle. This fossil is a very rare red-bed example of growth rings that might have been determined by moisture variance or abundance. The evidence of ring growth circa 305 million years ago is quite evident in this fossil which was located on the edge of what was once a wet plain with an extensive stand of ferns (Henry Ortiz, 2018).
Rainfall and Water Dynamics
of the El Pueblo Swamp
Top slab in above photo shows concave drip marks, which are rare. Drip marks (Henry Ortiz, 2014) formed from repetitive drip-drops falling from wet vegetation after a rainfall. Lower left and right raindrop-impressions are convex and concave respectively.
--------------------------------------------------
The rainfall dynamics of the El Pueblo ichnotelmafacies are both telling and interesting. The Great Spirit as the Natives referred to God has no equal when it comes to providing the proper elements for a living environment. First and foremost the El Pueblo swamp (swamp=the living environment, ichnotelmafacies=the present fossil quarry) was rainfall dependant. The marine environment and the water that supplied the marine basin was no longer accessible. Due to volcanic uplifts the land had risen high above sea level. Rainfall varied from light afternoon showers almost on a daily basis to occasional monsoon-like rainstorms that brought in a lot of hematite-reddish turbid water with lots of fine sand and clay. This floodwater flowed from swamp to swamp along the flowway, usually fed by narrow tributaries, that stretched all the way from the upland to the lowland. Rainfall and flooding represent every layer of sediment that concreted into layers of clay or rock in the El Pueblo swamp. The most common evidence of this rainfall and flooding is represented by raindrop-impression fossils and wavy or rippled rock layers. Two major records of raindrops exist in the facies. One is composed of concave raindrop impressions (actual raindrop-impact impressions) and the other is composed of convex cast impressions made by a succeeding layer of rainflood sediment that filled in the original rainfall impressions. (These "original or cast" distinction applies to all other trace fossil impressions including ripples, footprints and waves). Many people refer to these two distinctions, respectively, as a positive and a negative record. Professionals refer to these two distinctions as a concave (true impression or a convex (cast impression).
The water supply in the El Pueblo swamp varied from shallow to deep flood levels. However, because of its makeup the El Pueblo swamp was capable of draining excess water both through overflow and through regulated leakage.The El Pueblo swamp maintained both shallow and deep zones as well as vegetated shores that varied from shallow to deeper water. The design was to maintain hatching zones, swimming/hunting zones and variable vegetated zones where small animals could elude predators. Deep cool water at the center of the swamp served as recluse from the hot early Permian weather, which was tropical. Last but not least the El Pueblo swamp contained a high area and a high-shore side where small terrestrial animals could seek refuge during inundation.
Ethereal drainage of swamps, ponds, and lakes of the early Permian (a high evaporation rate) often substituted for lack of downward drainage. A humidity cloud around swamps, not unlike a rainforest, provided moisture for the intricate foliage of plants and trees.
Flagstone Quarries of the El Pueblo/San Miguel
Valley-bound Trajectories

Above photo shows amphibian footprints. The El Pueblo/San Miguel quarries were in operation for over fifty years. Known as San Miguel and Pueblo red, flagstone from these quarries attained a high popularity with stone setters and home owners alike. For quarry workers who did all the extraction work manually the flagstone demand nonetheless created much needed employment, even if at low wages. The work was hard but intriguing, often netting very large and very flat patio-paving stone as well as large thick slabs for outdoor benches. The thicker slabs were often broken into squares producing a picturesque medium for the construction of flower beds, retainer dry walls and border walls. Most of the quarries in the area are now exploited or impeded by the massive thicker strata that inevitably follows the extraction of quarry perimeter flagstone. Of all the quarries in the area the El Pueblo quarry netted the best surprise: a high number of footprints that predate the dawn of dinosaurs. Like the quarrying of quality flagstone the quarrying of good footprints entails a labor intensive process aimed at producing intact slabs with intact footprints (Updated as background info, Henry Ortiz early 2020).
Climate-Indicator Trace Fossil

Above photo: Finally, the traces of a climate indicator: A pincer trackway of a Fiddler-type crab of the early Permian (Uca Grandis, elpueblus). The space between the small and large pincer traces is about five inches, making this crab about fifteen inches wide, pods included. This crab identification is based on the fact that the claw tips are narrower than in a lobster. According to Dr. Ron Hines from 2nd chance info., ideal temperature for a Fiddler crab (anecdotally with one claw bigger than the other) is 75 to 85 degrees Fahrenheit. Any temperature below 65 degrees is detrimental to a Fiddler crab. A constancy of this temperature range can only be found in a wet-tropical climate, such as in a rain forest. It can therefore be inferred that the climate of the El Pueblo early Permian swamp was wet tropical, and that morning and afternoon showers were probably the norm. (This explains the repetitious presence of raindrop impressions throughout the facies.) Although daytime temperatures may have reached a hundred degrees or more, the cooling effect of regular rainfall kept the daytime temperatures at tolerable levels. Although there was a level of aridity a constancy of showers prevented desiccation. Dry-outs in intermittent swamps and flowways were quite brief, although concretion and mud-cracking could occur quite fast because of very hot daytime temperatures. The daytime evaporation rate was high. It was a one-season sort of environment except for one important interruption: monsoon seasons marked by a great influx of moisture from the main equatorial sea. A lot of dangerous flooding occurred amidst this great replenishment of moisture. Thank you, Mr. or Mrs. crab (might be a Mr.) of 285 million years ago, for this very valuable elucidation that finally sheds some light on the climate of the early Permian. Along with the footprints of very large dragonflies that I have discovered in the El Pueblo ichnotelmafacies the traces of a giant Fiddler crab further reinforce a wet-tropical climate. The predators for both genera were there, but so were the right climatic conditions for large growth. The great paleontologists of yesteryear correctly commented that the Permian and probably other Periods were but a terrestrial extension of the Carboniferous Period. That would mean that vegetation during the early Permian was quite lush.
The Saga of the El Pueblo Facies Continues
Above manus photo is that of Baropezia. The term Baropezia, which roughly means "heavy foot" first appears under Dawson, 1868. This footprint, along with many different others, was first attributed to the late Carboniferous Period. Slowly but surely the El Pueblo facies has matched many footprints from the late Carboniferous to the (Artinskian) early Permian. The above Baropezia ichno reference now becomes Baropezia el Pueblus (Henry Ortiz, 2019).
Other Crab Ichnotraces from the El
Above photo shows pincer and tip traces of an unknown specie of crab. The El Pueblo swamp facies has produced the walking traces of many different kinds of crabs as well as the traces of large scorpionoids. Parts of the swamp lane were obviously a marshland.
Above photo shows crab footprints (tip, spade, etc.). The El Pueblo facies is fast compiling an extensive and incredible number of same-site ichnofauna, availing more FYI for fossil buyers and researchers. Other than a giant Fiddler Crab trace, no other crab traces have been identified as to specific type.
The Trace of A Giant Arthropod
Above photo shows an under-impression of Arthropleura? (Meyer, 1884) which was a large herbivore arthropod. But wait. The absence of cross segment lines and a "soft belly" impression has thrown a monkey wrench into this identification. This might be a trace of a giant caterpillar-like insect, which nonetheless classifies as arthropodic. That would entail a tree-climbing caterpillar possibly capable of turning into a giant flying insect. The early Permian continues to hide many secrets, and deciphering them is no easy task. I have no choice but to revise this trace fossil as indeterminate between Arthropleura or a giant caterpillar (revised on 7/30/2019, H. Ortiz).
Rainfall From Nearby Seas


Photo on top left shows the form of a clay-buried probable fern frond from the v (vegetated) zone of the El Pueblo swamp. Ferns were soft, mostly non-resinous plants that were actually eatable, and that left very few impressions on redbeds because of ferric oxidation. In this instance the clay slowed down the oxidative process, disallowing quick dissolution of the vegetative matter. The photo on top right is a colorized version of the same plant. The fact that few fern traces have been found in the El Pueblo ichnotelmafacies belies the presence of lush and abundant vegetation during the early Permian. The reason vegetation was very plentiful was because there was no shortage of rainfall. This was due to the nature of continental drift and tectonics*. Continents are like icebergs, with greater mass below the surface of the ocean. As many continents slowly abutted and collided with each other during the formation of Pangea, plate subduction and volcanic upheaval occurred at a grand scale. Large inland-sea lakes were uplifted into mountains that continually weathered and kept uplifting at the same time. Because the actual collisions occurred deep below the surface of the ocean, continental slopes and shelves provided enough space for narrow seas to prevail between continents. These seas, as well as the uplifted terrain on which the El Pueblo swamp trajectory was located were coeval, and therefore of Artinskian age. For these narrow intra-Pangea seas and ocean bays, near-shore burial and volcanic gas/magma intrusions were common. This was a natural consequence of the uplifting process, which was ultimately caused by upward magmatic thrust and gaseous expulsions. The toll on marine invertebrates living in these narrow intra-Pangea seas remains unknown. Trilobites may have taken quite a hit. In line with the natural mandate of the day, which was the creation of variable elevations and terrestrial evolution, the paleo-tropical belt during the Permian was adequately supplied with rainfall. Rains were driven by ocean warming and distributed by equatorial-convection heat. Rains were often strong and brief because of fast-moving winds designed to broadcast precious moisture along or across the tropical belt. The large size of many tetrapods, some that weighed many hundreds of pounds; very large herbivores, as well as carnivores; all point to abundant biological mass. A truly amazing natural and Creative feat, yet one of great marine destruction in exchange for the terrestrial.
*Continental Drift Theory, Abraham Ortelius, 1596, and Alfred Wegener, 1912, from Wikipedia.
Photo above shows what may be giant fern spore-cases in clusters. Their spherical capsule-like shape may have enabled these sporangia to float and propagate on water. The duality of flowing water and drained areas during the early Permian called for a duality of propagation. These giant spore cases probably opened up through water saturation as well as through desiccation atop a moist substrate. Because only faint leaf-side current traces are evident (bottom of slab) a botanical identification remains indeterminate. The fact that the leaf matter left no trace points to a non-resinous soft plant, and to the scant ferric-redbed taphonomy of early Permian ferns. The round concave indentations were made by a modestly hard spore-case cover that did not crush until the form had been preserved.

Above: Three important types of tetrapods that lived in the El Pueblo early Permian swamp trajectory. The large amphibians had thick wide skulls with an unusually long mouth aperture. This allowed passive feeding by filtering small floating food, a must physiology for infant amphibians. Adult amphibians were powerful biters that could attack and kill other large amphibians and reptilians. The Pelycosaurs' temperature-regulating sail allowed such notable tetrapods as Dimetrodon to travel in the hot sun, and probably in cooler weather from one swamp to another searching for smaller prey. Some pelycosaurs were herbivores. The Araeoscelids were smaller thin lizards, either amphibian or reptilian, that could run fast during hunting and could avoid large predators through quick darting movements. It is logical that the first tree climbers were reptilian Araeoscelids. The illustration above (large amphibian) deals with that old question of stem amphibians. Stem amphibians were those little salamandroids from before the Carboniferous Period from whence the reptilians evolved. But during the early Permian, as elucidated by the El Pueblo swamp facies and other tracksites, the mega-amphibians (largest) were bigger than most of the mega-reptilians, if not all. Doesn't it stand to reason that during the early Permian the mega-amphibians were the stem amphibians because at various stages of early evolution the amphibians and the reptilians were roughly the same size, in paired genera, in order for survival of both. This conclusion is based on the fact that only after entire trajectories of evolution swamps disappeared did the mega-amphibians die off or became much smaller. The one characteristic that set mega-amphibians apart was their place in time, which foreshadowed the existence of small amphibious inland salamanders and frogs. The largest living salamander amphibians known today, upwards of five feet long, live in Japan and China. Environment was and remains key in the size of basic species which can vary in size according to food supply and environmental conditions.
A Bit of Chemistry From the El Pueblo Facies
Above photo: This Walchia Piniformis frond is outlined in hematite, no less. There was a great love affair between iron and oxygen in the El Pueblo swamp. Actually there was just too much iron in the water. But it was the iron that appropriated oxygen at the bottom of the swamp and delivered oxygen in the bloodstream of every tetrapod. Little has changed. The white splotches on the rock beneath the fossil are nothing less than petrified foam that contained suspended calcium carbonate. Copious amounts of calcium carbonate helped control the oxidation of the high iron content in the water. Nature's balancing act continues, and hard water is still present around the El Pueblo ichnotelmafacies.
A Gorgonopsid in the El Pueblo swamp?

Above photo: Gorgonopsid footprints, a very exciting, although sadly, a murderous hunting identification brought about by the discovery of a small manus (light purple color) and bolstered by the footprint reference: File:Orma di rettile gorgonopside, Aldino (Bolzano). Gorgonops remains are so far limited to south Africa (Richard Owen, 1876). This novel identification dawned on me while initiating the study of order of arrival of tetrapods into the El Pueblo swamp. Sure enough, Gorgonops was one of the very first tetrapods that wandered into the El Pueblo swamp, probably in search of small tetrapods brought in by floodwater. Gorgonops was a front-heavy barrel-chested tetrapod that was much lighter from the back. The large extended head and neck, which added to frontal weight, gave gorgonops greater distance between the deadly front claws and its large dagger-like fangs. Biarmosuchus, an animal very similar to Gorgonops that was found in Russia, has produced a constructive rendition that shows Gorgonop's front heavy body. The large manus and small pes suggests that Gorgonops was a fierce fighter and predator that stood its ground. These footprints from the El Pueblo facies comprise the first evidence placing Gorgonops in proto-North America. They precede other Gorgonops evidence found elsewhere by over twenty-five million years. The El Pueblo swamp facies has placed many ichnotypes within a coeval time frame, making earlier or later dates more inclusive. This latest Gorgonops footprint find shows that the sideways claw was not an anomaly, but rather a standard sideways claw for hunting or defense. The sideways claw on the manus is reminiscent of the lion's claw-hug when attacking large game, which inflicts claw injury to the prey, usually while delivering a paralyzing bite to the neck. Gorgonops has been equated with the saber-tooth tiger of the Permian. Eryops, the former designated Alpha of the El Pueblo swamp, is now in a different classification. Eryops was the Alpha of the amphibians, Gorgonops was the Alpha of the mammal-like reptilians. We now know that Dimetrodon elpueblus and Dimetrodon berea elpueblus, both considered accomplished hunters of small animals, were contemporaneous with one of earth's most fierce ancient creatures. Did Dimetrodon's thick skull and pointy sail provide a degree of deterrence and protection from a top-mounting assailant? If so, Dimetrodon was an ecological member, perhaps hunted, but not meant to be eradicated by Gorgonops. Gorgonops went on to fight deadly battles in the late Permian of Africa. And why not in the future north American continent if Pangea had not yet begun to break apart during the late Permian? (Henry Ortiz 2019)
Above photo shows the correct manus-pes arrangement of Gorgonops, the lion king of the early Permian and beyond. Although this is one muddy footprint-fossil it shows that the pes was much smaller than the manus. Nonetheless the pes digits were quite stout. The pes clearly left an impression of the "stabbing thump" that Gorgonops employed in its deadly hunts. A strong demand for this footprint, unfortunately, has been met with the fact that a Gorgonopsid footprint is so far the most rare footprint in the El Pueblo ichnotelmafacies. Only time will tell if more Gorgonopsid evidence becomes available.

Above photo shows an infantile footprint which finally makes sense. This is the manus footprint of an infantile Gorgonops which at an early age exhibits a dangerous gut-piercing spike. Measures 1-1/2 inches across. This is a soft-mud retraction footprint whereby mud collapse left only lineal impressions. Adult size manus footprint from the El Pueblo facies is seven inches long and was found at a flowway entrance into the swamp. Gorgonops means unbecoming but nature works in many ways: Gorgonop's thick skull (early Permian) was probably meant to deflect skull bites, a favorite fighting method of large predators, past and present. The two ichno-Dimetrodon species so far confirmed in the El Pueblo facies were of a smaller size, perhaps in the 75 to 100 pound range, maximum. So it was with the early Permian version of Gorgonops; this front-heavy tetrapod would have been much smaller than a middle or late Permian version. But Gorgonop's basic characteristics were already in place, of utmost significance and ecological importance, its tendency to hunt for large prey, inclusive of large herbivores, amphibians and reptilians. It goes without saying that the ecological web of early Permian life was self regulatory. That web of control was not any different from the savage carnal mechanics of modern wildlife that co-exists in the interest of maintaining, but not destroying, the habitats and all of the inhabitants. A balance of diverse fauna in the right ratio to the floric environment, was a prerequisite meant to further terrestrial evolution. The system of protoplasmic protein exchange, which many of us grumble about even while we eat meat, continues onward, creating in its wake, many a vegetarian, and perhaps an evolutionary awareness toward the more spiritual. Yet, the fact remains that for humans, proper amounts of animal proteins and carbohydrates from infancy to old age remain essential for health and longevity.
 |
| The area of the El Pueblo swamp facies is about three acres |
Not to worry! The El Pueblo early Permian swamp facies (facies means a particular rock face) was a flagstone quarry that ceased operations because of its rich trace fossileria. At the present only small investigative excavations are being carried out. At least eighty-five percent remains intact and will be left that way. Attempts are underway for placement of site under conservator status. This is the site that elucidated Ichniotherium cottae's ground-sloth-like identity and also defined for the first time the geological, hydrological and hydraulic mechanics of early Permian evolution swamps.

Update:
The El Pueblo early Permian swamp facies has finally completed an ecologically viable count of tetrapod footprints. Coupled with plant impressions, myriapoda and insect trackways, the El Pueblo facies now represents one of the best non-marine ichnological quarries in the entire world. The site has surpassed the ichnotetrataxa count of amphibian and reptilian footprints from any other single source. As many have expressed, it's the paleo site that is the real teacher. The El Pueblo facies established a world-standard as to the ichnotaxa variety of the early Permian. The support flowways exhibit a much smaller ichnotype group, which although related to the El Pueblo facies suggests who the more transient tetrapods (or often, water-stranded amphibians) were. Some tetrapods were dwelling in deeper and bigger flowway-swamps with a higher permanence of water supply than in the El Pueblo swamp. Those habitats were not leaky swamps with an aggradation of concreting layers, therefore the footprints are harder to find.
Fast Tetrapods of the early Permian
Many tetrapods from the early Permian were fast and agile. The photo above shows a running-context trackway of a probable araeoscelid tetrapod chasing prey or escaping from another predator across soft mud. Running-context trackways are very common in the El Pueblo early Permian swamp facies. Other smaller footprints on slab are from amphibians.

Above photo: Footprints of two lizards scampering across a semi-hard mud surface. Probably araeoscelid hunters.
Savagery of the Ages: Two
Flowway Hunters Identified
A.
B.
 |
A true early Permian crocodilian footprint
about four inches across. |
C.

Above photos show the footprints of early Permian flowway hunters. The EL Pueblo swamp was fed and drained by flowways. The term flowway is used in El Pueblo ichnotelmafacies research as an alternative to rivers because of their intermittent nature. The investigation of the Agua Verde (green water) and Encinitos (little oaks) flowways near the El Pueblo ichnotelmafacies revealed two common footprints of probable carnivore-reptilian entity. Photo (A) shows the infantile footprint of a probable crocodilian-like reptilian in a shallow-water swimming context. Note three digit manus that might be reflective of the later crurotarsal dinosaurs. Adult size, about three to four inches wide, of same ichnotype crocodylomorphs have been identified in both the Agua Verde and Encinitos flowways. Photo (B) shows the manus footprint of a true crocodilian with four straight short digits and claws. (C) shows the pes footprint of Dimetrodon berea (Tilton, 1931), at about five inches long. Both reptilians were inclined to wait for prey, although the straight claws on crocodylomorphs also enabled direct attack. The blade-like impressions of Dimetrodon berea's claws now suggest an improved claw set (hybrid claws) designed for both static and attack traction. What better place to hunt or waylay prey than within a deep flowway-swamp full of young Dromopus-species tetrapods.

Above photo: One of the neatest things I have noticed about Dromopus Marsh is the fact that these were "herd animals". The behavioral context for the above group of footprints, modernly called a lounge of lizards, is directional unison. Perhaps seeking safety in numbers, large groups of Dromopus Marsh traveling in the same direction must have been a sight to behold. Directional unison is apparent in recently hatched broods as well as in larger track makers. Adult footprints seem to be more singular, and there is that probability that the Dromopus sp. group was susceptible to a high level of predation. Dromopus footprints are common in the El Pueblo facies as well as in related flowways. Slab is nine inches across.

Above photo shows the trackway of a dragonfly. The undulating line is a mud crack. Using its long back appendage this particular dragonfly was laying its eggs in very shallow water. Other dragonfly trackways from the El Pueblo facies also show mud cracks ideal for depositing eggs so that the hatchlings, called nymphs, would remain localized. Slab is eleven inches at bottom edge. This dragonfly was sizable. Trackway width to wing-span ratio means this dragonfly was sporting a ten-inch wingspan. Wider trackways of unknown species of dragonflies have been found in the El Pueblo facies.

Above photo: The size of this insect (revised to a Griffin fly) was absolutely not funny. Some say it would have merely boxed your ears with its wings, but then what about its bite? Meganeura and Griffinflies are different term that refer to the same very large dragonflies of the late Carboniferous Period. Talk about a good food and oxygen supply. The footprints alongside ruler by the number 1 and the number 7, and another one to the left, seem to include feelers (vibration sensing organs). This particular ambulant was taking four-and-a-half inch steps The footsteps might very well be quadrupedal. Unknowns aside, giant insects are a clue to a rain-forest type of environment. The width of the dragonfly trackway across the middle length of photo represents a dragonfly with at least a twelve inch wingspan. The trackway with the four-inch footsteps probably represents a dragonfly with a much wider wingspan that matches the El Pueblo facies' age of 305 million year ago.

Above photo: Closeup of Griffin fly/Meganeura insect footprint reveals three side-ward hooks. A Griffin fly was an insect that was relatively heavy and the hooked claws were probably used for quick perching, probably sideways, from fern or conifer trees, etc. Lineal impressions represent long feelers which probably had hairs. They were also pollen dusters that picked up pollen from plants and from moist mud. The pollen was transferred to other plants and or dusted along moist mud. There seems to be a fourth side-ward hook by broken edge of fossil.
These are probably the footprints of the famous Griffin fly, Meganeuropsis (Carpenter 1939), of Pennsylvanian age. Griffin flies sported a wing span of up to 28 inches. With an age not too distant from the end-Carboniferous Period, the El Pueblo swamp was a place where swimming swamp creatures interacted below flying or tree-perched dragon flies that were very big and colorful. It is probable that dragon flies flew from swamp to swamp along the swamp trajectory, which extended all the way to the upland.
Other Significant Fossils from the El
Pueblo Early Permian Facies
 |
| Dimetrodon was named by E. Cope, 1884 |
Above: Dimetrodon elpueblus footprint. Note the human-like plantar region that was characteristic of large pelycosaurs. A long plantar region allowed for rearing on hind feet for attack or defense. This Dimetrodon pes footprint has straight-aligned claw ends, perhaps meant for better traction. Visit the blog "Dimetrodon's Ichnology" for more Dimetrodon footprints from the El Pueblo ichnotelmafacies.
A Guinea Pig-like Footprint
 |
| Espuelacerta (Henry Ortiz, 2017) |
 |
Supportive fossil evidence
for Espuelacerta, 3 inch long |
Above: Was there a tetrapod with a downward heel spur in the El Pueblo early Permian swamp? I had determined that only the discovery of other similar footprints would confirm that theory. Luckily most discoveries are made by accident and by accident I found two other similar footprints. So apparently there was a Permian tetrapod in the El Pueblo facies that possessed a heel spur for traction. Evolution swamps of the Pennsylvanian/early Permian, which had a lot of clay, were muddy and slippery environments. (Please note that all new ichnological names from the El Pueblo facies include the suffix elpueblus for locality, e,g., Espuelacerta elpueblus Henry Ortiz 2017.)
Little by Little the El Pueblo Ichnotelmafacies is
Revealing More Previously Undiscovered Footprints
Above photo: Tentative assigned ichnological name for above footprint set is Duoespuelacerta elpueblus (Henry Ortiz, 2017). There was a need for traction spurs in the beginning early Permian as is exemplified in the above footprint set. The manus is tridigit but with two side spurs. This footprint might belong to the Pennsylvanian amphibian Megabaropus hainesi (Carmen 1927) if same discoverer missed the traction spurs. There is also the possibility that the traction spurs developed during the transition from the Pennsylvanian inland marine to the early Permian swamp environment, in which case the ichnological name Duoespuelacerta elpueblus, an amphibian, becomes definitive. The pes is quadrudigit with spur-like, downward-tipped digit ends. Three and four-digit combinations were common during the end Pennsylvanian and neo-Permian. Pes is five inches long.
Above photo shows a slightly enlarged five-digit footprint of an indeterminate tetrapod. The El Pueblo facies exhibits a very high ratio of single or double footprints in comparison to the number of footprints in trackways. Note the hand-like configuration and what appear to be bifurcate claws.
Above photo: A planter region as round as a ball. This might be the manus footprint of Ichniotherium cottae. The roundness of the plantar region was both mud-sink resistant and swiveled well on soft and hard mud during each opposite-side step or because of limited limb radius. But just as important, this ball-like plantar region added traction in soft clayey mud.

Above: The only conclusive evidence of a webbed foot so far. Webbed feet were common during the early Permian. Hopefully the El Pueblo early Permian-swamp facies will provide other fossils with evidence of webbed feet. Often the clue to webbed feet are very thin but unjointed digits.
Trackways from the El Pueblo Swamp Facies
 |
| Enlarged at crisscrossed point. Footprints slightly larger than in photo below |
The above trackways are probably those of Saurichnites salamandroides, meaning footprints of lizard-like salamander (Geinitz 1861).

Above photo: This trackway from the El Pueblo facies matches perfectly with those found in Europe. Some paleontologists attribute it to Batrachichnus delicatulus. The context of the trackway is difficult to distinguish from distressed or hurried. The wide occurrence of this type of trackway, which is common in the El Pueblo ichnotelmafacies and in Europe, speaks volumes about the universality of the early Permian, including facies and taphonomy. This type of trackway is a good guide fossil to the identification of an aggregate environment bearing the remarkable universal geological mechanics of an early Permian metropolis and biofacies. The track maker Batrachichnus delicatulus exhibits evenly spaced pes and manus footprints, rather than sets of pes/manus footprints. Yet on closer examination as in photo below, smaller footprints are visible alongside the larger ones.

Above photo is a distressed context trackway of Batrachichnus delicatulus. The tail-drag mark and high relief convex cast footprints mean this tetrapod was pretty well bogged down in soft mud, making it prone to predation.
Photo above shows even-pace footprints of a probable Batrachichnus. But in mobile ichnology things are not that simple. Although footprints at even intervals are one of the Batrachichnus interpretations, this trackway shows that pes-manus sets become problematical when an animal was scampering or otherwise moving fast, or even when being held back by mud. The two preceding photos also show footprints where pace and stride become irrelevant. The ichnology of running and walking footprints are worlds apart, especially when the diagrammatic mapping is based on digit count alone. The "mud tracking" tetrapods were quite numerous.
Update on Batrachichnus Footprints
from the El Pueblo Swamp Facies

Above photo: Batrachichnus simply means frog or toad-like digits/ footprints. These footprints appear to be four-digit (tetradactyl). They can be attributed to Batrachichnus plainvillensis (Woodworth) insofar as four digits. Could be Batrachichnus but as the saying now goes (based on so many variations and sizes of Batrachichnus footprints): I just can't blame everything on Batrachichnus delicatulus or salamandroides. The term has become so generalized and overused that its application to so many different footprints sometimes threads on vague. During the early Permian the variety of salamander-like and frog-like amphibians was extensive and needs to be specified further based on subtle ichnological differences and stride and pace that depart from any frog-like manner of ambulation. The question is: Were all four-digit tetrapods of the late Carboniferous and early Permian one family or do four-digit footprints represent a root clade of the Erypoids and other four-digit temnospondyls? If so, that would explain the many large four-digit footprints in the El Pueblo ichnotelmafacies. An interesting parallel of footprints was recently discovered in the El Pueblo facies: tetradigit and tridigit footprints are present in an area that contained a lot of mud. This is in line with my early theory that less digits constituted a design originally geared to the shedding of mud. Three and four-digit footprints usually radiate from a sink-resistant plantar region of adequate size. Another question under investigation is the presence of large four-digit footprints (with large plantar regions) alongside the smaller Batrachichnus plainvillensis footprints. The smaller footprints might be infantile, not adult I'm at a loss as to when real Pennsylvanian frog jumping-stride footprints, with even side by side pes and manus, will emerge in the El Pueblo facies.(Henry Ortiz 2020).

Above photo shows tetrapod footprints in watery clayey mud. This is a trackway of a typical small tetrapod headed in the direction of deeper water because exposure of smaller animals was a danger point in a Permian swamp. This quagmire usually happened when the same flood water that brought smaller tetrapods into the swamp drained rapidly through overflow, leaving the new arrivals stranded on watery mud. One of the facets of the El Pueblo and other early Permian swamps was that rain-runoff episodes created a live-food trap. The likes of Dimetrodon and Gorgonops surely looked forward to this muddy episodes. This post-flood feeding mechanism still exists along rivers. Holistically, the prey-predator mechanics within early Permian swamps and earlier represent well what would be sustained in life to the very present. The only noticeable difference is a modern growing attempt attempt at vegetarianism (which is detrimental to infants) among the human species.
Ichnological Examples From the
El Pueblo Facies
A.
B.
Above photo (B): Classic Anthichnium footprint, about 1-1/4 inch across. Anthichnium, meaning flower-like footprints, Pabst, 1900, often left a tail-drag impression. (A) shows a partial trackway with a tail-drag mark which nonetheless looks normal enough. But Anthichnium was a five by five-digit tetrapod (not four by five-digit [a variant?]). Judging from the undulation of the tail drag, Anthichnium was an excellent swimmer. Its profile was low, typical of those side-limbed smaller tetrapods able to recluse in shallow water. Once again the common salamander becomes the bearing point for an identification. What else is available for comparing amphibious footprints but frogs and salamanders. An adult Anthichnium was about eighteen inches long. Its footprints are popular with collectors because of an undulating nature and relatively large feet.

Above photo: Were there tetrapods capable of bipedal ambulation during the early Permian? Chances are, yes. Bipedal footprints exist in the El Pueblo facies. But some "bipedal footprints are sometimes deceiving. The above footprints seem like a likely bipedal trackway but the wavy texture suggests flowing water. This particular tetrapod was probably swimming in shallow water and touching the bottom with its pes while doing so. Because the tail-end of tetrapods was usually heavier, and the hind limbs longer, it is possible that the pes were touching the bottom while the manus were paddling clear off the mud. One thing is certain: tetrapods of the early Permian had retained the undulation movements of their fish ancestors and undulating their submerged tails in water of ample depth, were excellent swimmers. Digit impressions average one-and-a-half inches long.

Above photo: These footprints are often referred to as those of Limnopus sp. (Marsh 1894), and at one time were attributed by some investigators as belonging to Limnopus vagus. The footprints appear four-digit and therefore Limnopus sp. remains the safest identification. Limno simply means pond or lake and during the early Permian most feet were geared to muddy environments. Limnopus footprints sometimes exhibit digit-end curvature as in footprints of Anthichnium, and sometimes the digit impressions are evenly spaced and straight. The differential is probably a sideward stress variable as Limnopus scampered sideways or turned in a different direction (or) a response to muddy-soft or hard-dry surfaces. I have not yet been able to determine if the digit-ends are blunt claws, callused ends, or if Limnopus sp. possessed digit ends that served as traction pads in the manner of the gecko. I too will call upon modern frogs, salamanders, and geckos and favor the latter.
The True Hyloidichnus Bifurcatus
The name Hyloidichnus bifurcatus has been applied to a lot of early Permian footprints, including long-claw marks that merely suggest a true entity. The definitive footprint of Hyloidichnus bifurcatus has finally surfaced in the El Pueblo early Permian swamp facies. The true Hyloidichnus bifurcatus (Gilmore, 1927), was a sizable tetrapod. Its five digits seem to have been quite flexible and at the end of these digits was a characteristic that set it apart from many other tetrapods of the early Permian. That one characteristic, as the suffix "bifurcatus" suggests, is a split claw at the end of each digit. "Forked claws" surely had some advantage for Hyloidichnus, a utility now revised as for static traction on slippery mud (Henry Ortiz 2017). Photo (A) below shows the split claw marks of an indeterminate Dromopus-like ichnotype just above edge of white slab, and which shows the need for an encompassing genera or class of a "bifurcatus" group). The white slab is a classic representation footprint of Hyloidichnus bifurcatus.
A.
Grouping Bifurcated-Claw Footprints
As a rock-quarry laborer turned accidental early Permian podologist, the details of many footprints never fail to amaze me. Or to confuse me. Not too long ago I realized that the bifurcate claws were standard in several unknown ichnospecies. The footprint below exhibits the forked claws of the classic ichnotype Hyloidichnus bifurcatus. The footprints that follow C,D and E, exhibit the forked-claw mud-braking configuration that was more common than previously thought (Henry Ortiz, 2017). (Please note that self-cites, which already sound redundant, are provided for matter of record only, and allude more to the site than to discovery.)
B
 |
| B. The true Hyloidichnus bifurcatus |
Photo (B) above shows the full length of Hyloidichnus bifurcatus's pes footprint at about 2-1/2 inches, probably adult size. The general physiology of the plantar region is roundish, with digits (some missing) fanning out.

Above photo: These are the footprints of Dimetrodon berea (Tilton, 1931). The fact that Berea, West Virginia and Ribera, New Mexico are over 1500 miles apart further reinforces the universality of early Permian fauna. Dimetrodon berea possessed bifurcate pes claw ends in contrast to the classic Dimetrodon that possessed straight claws. Because bifurcate claw-ends are designed for static traction this led to the supposition that Dimetrodon berea was a waylay hunter. But a revision is in order (8/7/2018). Dimetrodon berea's footprints exhibit (via better footprint specimens found lately) sideward-curved claws on manus and bifurcate claws on pes. Dimetrodon berea's narrow trackway suggest a narrow body like other Dimetrodons. However, that narrow trackway could increase in width everytime a Dimetrodon walked in a more sprawling posture. But why curved claws on manus that resemble those of the herbivore? If Dimetrodon berea was a reptilian predator it is probable that it hunted both in the water and in vegetated areas. Because Dimetrodon berea was primarily a flowway hunter, and not primarily a swamp hunter like Dimetrodon elpueblus, it probably hunted in protracted vegetated shores. However, we remain confronted by that curved-claw peculiarity that was designed to minimize upward pressure on the phalanges when walking on hard or concreted surfaces and to avoid hooking on vegetation . The big question remains: Were many tetrapods of the early Permian able to position side-curved curved claws into a more erect position for traction, or while in attack or defense mode, or while digging through humus layers? A physiology of such duality would have entailed an unusual degree of digit joint radius normally accomplished by double-bone limbs, as from the wrist to the elbow in humans. Or was it the wrist that was swiveled sideways in order to place sideward-curved claws in a more operable position for clawing? Either way, the design worked on the many tetrapods with long curved claws in the El Pueblo swamp.
A.
Photo (A): Dimetrodon berea's manus (might be pes) in action. Note how deeply the manus was anchored into the mud and how two of the bifurcated claws (toward right) seem almost T-shaped in order to maximize static (stationary) traction. There is another concern in all of this: If Dimetrodon berea's bifurcated claw ends were blunt at the top and blade-like at the bottom then we might be looking at the hybrid claw-ends of the Pennsylvanian/early Permian. Dimetrodon berea's bifurcated claw ends might have increased traction when fleeing from other predators or while attacking prey across a muddy surface. The above footprint might be a mud compressed trace of the pes, which would make sense for faster predatory attack via bifurcate claws that were blade-like at the bottom.
B.
Photo (B): In this partial trackway of Dimetrodon berea, which is narrow and suggestive of erect limbs (H. Ortiz, 2017), the bifurcated claws barely recorded. Dimetrodon berea's claws were probably well elevated for normal ambulation and only when hunting in soft mud were the feet and claws oriented downward allowing for static traction.
C.
Photo (A) and photo (C) suggest that Dimetrodon berea's bifurcate claws were blunt at the top and blade-like at the bottom. This ichnological design may have enabled D. berea to hold down prey and to dig in leaf mats. The T-blade-like claw ends were also appropriate for easy stabbing into mud or moist ground.
 |
D. Pes-footprint length 3" long.
|
Initially confused with Hyloidichnus bifurcatus, above footprint, with a more elongated heel, remains indeterminate. More and more the El Pueblo swamp Hyloidichnus bifurcatus (Pennsylvanian) seems to have been a tri-digit forebear or another specie.
 |
| E. Procolophonichnium ispp., 7" long |
Procolophonichnium ispp. (Nopcsa,1923), was known only from the Triassic before surfacing in the El Pueblo early Permian swamp facies. This is the footprint of a very large reptilian "mud hunter" with blunt claw tips that also exhibit blade-like bottoms. More details in following subject, Large Tetrapods of the Early Permian.
A Surprising Conclusion to Hyloidichnus
bifurcatus and Other Bifurcated Claw Tips


Above photo: When Charles Whitney Gilmore named Hyloidichnus bifurcatus he was probably referring to the hyoid (u-shaped) bone in reference to the V-like claw tips. A new find in the El Pueblo early Permian swamp facies has shed new light on the merits of Hyloidichnus's odd claw tips. The above claw impressions might belong to Dimetrodon berea. They were not odd at all, but rather a manifestation of functional design fit for the time and for Hyloidichnus's hunting habit. Since a long time ago Hyloidichnus's footprints, first reference to bifurcation, in the El Pueblo swamp facies have suggested hunting or foraging inside small streams or drainage paths, and some footprints have suggested that Hyloidichnus bifurcatus waited for its prey. This tetrapod, along with its large stubby clawed cousin Procolophonichnium ispp., Dimetrodon berea, and other bifurcate ichnotypes lay in wait, although it is not clear if these tetrapods were herbivore, carnivore, or both. Nonetheless forked blunt claw ends (shown above in a stopping context) were not claws as much as an anchoring or a braking device--the proper mechanical feature to keep steady inside a rivulet or on a swamp slope in wait for flowing organic matter or while feeding on vegetation. The muddy flowway and swamp environments of the early Permian led to the evolution of blunt, bifurcated claw ends, which are proving to be more common--and functionally complicated--than previously thought. (Henry Ortiz, 2017.)
Stenichnus Footprints
A.
B.
Above photos: Footprint of the amphibian Stenichnus, first discovered by Gilmore in 1927. Stenichnus means long straight digits. Photo (A) shows a Stenichnus pes with bifurcate claws. One digit on manus also suggests a bifurcate claw. Footprints suggest both a digging and a slope feeder physiology. Manus impression is two-and-a-half inches long.
Stenichnus and Dimetrodon Footprints
on Same Slab
Above photo shows a Dimetrodon pes sharing the same mud surface with Stenichnus (same slab as A and B above). The thick skulls of Dimetrodon displayed in some museums suggests that Dimetrodon was a powerful biter and bone crusher. Dimetrodon was probably shunned by others more than by its own kind. This proximity of a Dimetrodon and an amphibian footprint seems worrisome, although it is suspected that Dimetrodon preferred small animals, especially amphibious offspring. This reminds me of a line I once read: "If by chance we pass each other somewhere, do not forget that we are on different paths." Perhaps either the Dimetrodon or the Stenichnus set foot first, safely before the other walked by. The natural instinct of any amphibian was probably to steer clear of Dimetrodon.
Swamps Will Be Swamps
 |
| Left: Permiaphytofloter Ortizii, Henry Ortiz 2015 |
Although conifers and ferns are the signature flora of the early Permian the above two fossils might represent aquatic additions. The fossil on the left might be part of a floating or semi-floating plant, perhaps a swamp lily of the early Permian. The fossil on the right exhibits tapering ends of some sort of swamp plant such as semi-aquatic rushes. Only additional fossils will confirm the validity of these two plant fossils. Squares on background are one square inch.
Intertwined or Twirled Vegetation of the Permian Period: The answer might be written in the wind and in water currents.
 |
| Phytofunem ElPueblus, rope-like plant, Henry Ortiz, 2017 |
Above: Around an indeterminate footprint can be seen the post-flood debris of an early Permian monsoon rain. This particular debris, which was not debris in the first place, served its purpose as organic supply for the engenderment of swamp plankton. These are bits of an unknown plant with stems and or branches that were intertwined and that had the appearance of rope. When plants are rolled about by water currents or twisted by the wind, that old adage comes to mind: better to bend (or to give) than to break. Only a flood of runoff water could shred these plants into bits.These are remnants of dessicated vines, previously undiscovered in other facies.
Above photo: Other examples (thin one recent) of rope-like plant or vine or whatever this twirled plant-stem can be called. If a vine, this was the tree-climbing facility for insects and possibly a small-animal link between trees. Vegetation of the El Pueblo early Permian swamp was quite varied and hints of exotica.
Photo above shows what might be a plant stem with lenticels. Lenticels in tree bark are modernly widespread and allow gas exchange with the atmosphere. Because of a high level of carbon dioxide from volcanic venting during the early Permian, lenticel bark, if that's what the fossil represents, may have existed in more abundance than previously thought. The stem is of convex form.
Indeterminate Perfectly Round
Light-colored Spots
A.
B.
Above: For a hundred years flagstone-quarry workers have been calling these perfectly round spots on flagstones "moons". These spots are common on flagstones from flowways of the early Permian. The light spots are actually oxidation traces where oxygen from vegetative matter expended or oxidized the ferrous tint. The spherical shape of the vegetative matter points to large and small spore cases being distributed downstream by water flow, and that became trapped in sediment. The spore cases might be from very large ferns, but a definite identification is still lacking. Photo (A) exhibits a small and a large oxidation spot. Photo (B) shows cross sections exhibiting spherical shapes. Both wind and water current were important vehicles of floric dissemination during the early Permian.
An Important Discovery
White areas, splotches, fern forms, etc., on red rock have led to a very interesting discovery. It turns out that the redbeds of the El Pueblo facies, and many other redbeds, are actually light-colored quartz-sand beds. The red color is the result of a pink feldspar and hematite pigmentation. The discovery is not the above, but rather the fact that oxidative differentiation and reduction prove the potential of fluvial inclusion or exclusion of pigmentation during the early Permian, and that the inclusion or exclusion of iron, respectively, made the difference between biological richness or impoverishment. This reddish pigmentation often percolated downward or oxidized away, leaving the shapes of organic matter. Thus, quartzitic white beds atop the mesas around the El Pueblo facies, that have for so long confused geologists, are from granitic quartz, the same as the "redbeds" of El Pueblo facies. In Permian times iron became diverted along the alluvial clay valley, hence dinosaur skeletons of the Triassic are found along rivers. Also, highly hydrated iron as limonite is present on the white quartzitic beds, pointing to some level of fertility. Nonetheless, the hematite redbeds of the valley bear the true evidence of early life. (From the study, Investigating a Small Islet of the El Pueblo Early Permian Swamp facies (Henry Ortiz 2020.)
Tridactyl Footprints of the Early Permian
So why is it that dinosaurs, which were descendants of early Permian quadrupedal tetrapods, developed a tridactyl pes? It is obvious that nature decided that a three-digit pes was more suitable for a bipedal posture in many types of dinosaurs. Swamps and lakes of the Mesozoic were muddy affairs and the more digits, the more mud that could adhere to the primordial foot. Bipedalism itself hints of a conscious desire for height which provided vantage point when looking around for food. Bipedalism also allowed wading in deeper water. Apparently all tridactyl dinosaurs were carnivorous or omnivorous and were also able to run very fast after quarry. An ostrich can run at about forty-two miles an hour, maximum. Top human speed is 28 miles an hour. So even if dinosaurs could not run as fast as an ostrich, even at thirty-three miles an hour a modern human would have become prey on open terrain. The exact beginnings of the dinosaur tridactyl foot is unknown but its rudimentary form surfaces every now and then in early Permian swamps. Although the El Pueblo swamp facies has not produced any tridactyl footprints provably connected to the classic tridactyl dinosaur footprint, a three digit physiology was assuredly present. We must keep in mind that guinea pigs, birds, and other animals are tridactyl and so the three-digit physiology continued even after the dinosaurs became extinct. Below are two three-digit examples from the El Pueblo early Permian swamp facies.
 |
| Unknown and unnamed ichnotype |
Above: This is probably the footprint of a webbed tridactyl pes. Notice the heel spur on the back. A heel spur became standard in the tridactyl pes of bipedal dinosaurs. Squares represent one square inch.
 |
| Trigeckodactylus (Henry Ortiz 2016) |
Above: These three-digit pes and manus footprints apparently belonged to an amphibian. The pes footprint (back) exhibits what might be calloused-rounded digit ends for better traction, and perhaps for standing up on two feet. Although the exact details of foot evolution from the Permian to the Triassic remain unclear, the functional form of the tridactyl foot was alive and well in the El Pueblo early Permian swampsite.
Unidentified Footprints
The footprints below were discovered in the El Pueblo facies in 2017. All three are the same type. Perhaps they were made by a tortoise but more evidence is needed before that identification is confirmed. (Update, 2019): A strong possibility comes to mind: Could these be the first Artiodactyla (cloven-foot animal) footprints? They are not very common in the El Pueblo swamp facies suggesting that these footprints were made by roving tetrapod feeders not restricted to a swamp environment. They were quarried at the perimeters of the facies. The cloven foot tends to distribute weight evenly on the heel and hooves. The hooves were thick and sturdy and above all, wear and abrasion-resistant as is appropriate for walking on hard ground or on bedrock.
 |
| Ususduodactylus (Henry Ortiz, 2017) |
Large Tetrapods of the Early Permian
Tetrapods were lizard-like animals that existed millions of years before the dinosaurs. They were descendants of the first lobe-finned fishes whose fins evolved into feet. The first walking fishes eventually became terrestrial, able to breath freely out of water. The footprints of tetrapods have been found in large numbers in tracksites around the world. Their reproduction rate was very high, especially among amphibians. But footprints of the adult mamas and papas are found only occasionally. So how big were the larger tetrapods of the early Permian that laid eggs in shallow ponds and swamps of 285 million years ago? Very few skeletons of large amphibian tetrapods have been found. (Please note that not a single skeleton of any type has be found in the El Pueblo swamp facies.) The prominent amphibians were larger than the reptilians. Some measured over six feet long. But the largest skeletons of early Permian amphibians have yet to be found. Some adult amphibians weighed as much as three hundred pounds or more. Many large footprints by which to ponder have been discovered across the world and here are three local finds. One remains indeterminate until further correlation. These footprints (below) are testimony that the landscape of the early Permian was occupied by conspicuously large amphibians. Large enough to provide a conspicuous presence while walking around swamps. Their color? A rough estimation can be made based on nature's camouflaging colors: Reddish hues to blend in with the soil, greenish to blend in with the vegetation and or grayish to blend with the shade of ferns or deep water.

Large (seven inch) footprint with visible ball-like plantar region and long slender digits. Digit ends exhibit wide claws for good traction. This guy was a real mud walker and possibly an amphibian. Double indentation of one claw tip indicates overstep of manus digit (correction from fidgeting). Might be a large version of Procolophonichnium ispp. This rare type of footprint remains without any definite ichnoidentity and has been described as a parareptilian (Baird, 1984).
 |
Ichnogenus Eryops elPuebus manus imprint. Eryops was
the Alpha, the amphibian carnivore killer, of the El Pueblo
early Permian swamp. |
|
|
Eryops elPueblus pes, 7-1/4 inch long.
|
Above two photos: So what's with Eryop's four-digit manus and pes physiology? Ichniological evidence from the EPEPSF points to three and four digits in most amphibians. The digits often exhibit a play of side-ward flexibility if slender (as in Dromopus and other yet unidentified ichnotypes), or a blunt straight appearance if the digits are thick. Apparently Eryop's digits were of the latter type because of its considerable weight and the need for mud-sink resistance. In contrast to Dimetrodon's five digits on both pes and manus, the answer is quite clear. The more digits in a carnivore reptilian the more claws for holding and injuring the prey or enemy. Eryops was more of a mud and water dweller, and as such, depended more an a large wide mouth that usually waited for its prey, especially prey that was targeting its offspring. A probable omnivore, Eryop's feeding method stems from its infantile nature. Newly hatched Eryopods were passive feeders, hence their wide mouth which they held open in order to trap flowing eatable matter. Their large plantar regions and thick stubby digits kept them from sinking in deep mud while doing so.
A.
B.
C.
Above photos. (A): Barely an inch wide, this is a photo of an infantile ichno-Eryops elpueblus. Two things stand out in this footprint: the high relief means that the concave mold was deep and the product of a tetrapod that was unusually heavy relative to its size. The overstep of the pes means that Eryops elpueblus was a tallish high-stepper and not a sprawled walker as previously thought. Eryops elpueblus was actually the heavy-weight of the El Pueblo Artinskian early Permian swampfacies. (B): Another infantile Eryops manus footprint next to that of a probable (or similar) infantile Ichniotherium cottae footprint. These two track makers were probably not at odds since Ichniotherium was an herbivore and Eryops was a carnivore that probably fed in different environments. Both were subject to the slippery mud of swamps. For traction Eryops relied on thick straight digits and claws while Ichniotherium cottae possessed a propeller-like ring of side curved claws and a broad sole, And was the extra claw in back of this baby "marking animal" footprint (under penny) meant to increase traction? Why not, grazing on a muddy slope demanded as many traction devices as possible. (C): The manus footprint of Eryops elpueblus not fully grown. Shows wrinkles that might be a skin trace.
 |
| This pes footprint is identified as that of Edaphosaurus, an herbivore. |
(B)

Above photos: These footprints from the El Pueblo swamp facies resemble each other based on one of the digits being very short in comparison to the others. Both (A) and (B) have been revised to pes footprints and tentatively named Ichnoligodigitus (small digit) Elpueblus for reference. Note that many of these footprints might belong to well-known tetrapods. Without skeletal reference it remains impossible to assign some footprints to known tetrapods. (Because skeletons are usually found in an incomplete or highly scattered state, most of the reconstructed pedes and manus are estimations.) Footprint (A) exhibits what might be side-curved claw-end impressions. Side-curved claws often signal an herbivore entity. An amphibious designation remains likely (but not definite) because the muddy environments of the early Permian demanded claws for large amphibians that often walked rather than swam because of shallow water, including across or along muddy and sloped swamp shores. It must be kept in mind that the biggest early Permian herbivores, Ichniotherium cottae and Edaphosaurus, were reptiliomorphs. The differential here might be that some herbivores were roving reptilians while other large herbivores were swamp-bound amphibians precisely assigned to trim swamps that became overgrown with flora.
 |
| Possible amphibian mom and offspring manus footprint. |
Above photo: This pes footprint from the El Pueblo facies matches the amphibian Megabaropus hainesi (Carman, 1927), discovered in the Pennsylvanian of Ohio. Length of footprint at longest point is five inches.
Above photo shows an EPEPSF footprint matching Psuedobradypus ohioensis, a large amphibian, discovered in the Pennsylvanian of Ohio by Carman in 1927. In relation to their Pennsylvanian designation both Megabaropus and Psuedobradypus adhere to the (Artinskian) designation of the El Pueblo early Permian swampfacies, which is relatively near in geological time to the end-Pennsylvanian Period. Above pes footprint measures six inches across digit spread.
Kudos to the Early Amphibians
Early amphibians, such as Eryops and others, are usually referred to as primitive amphibians. Primitive amphibians ranged in size from salamander size to huge heavyweights that out-sized and out-powered large land-roving reptilians. This virtual army of amphibians that had a geological range from the Carboniferous to the Jurassic Period had something in common: they were more swamp bound and swamp proximate than advanced carnivore reptilians. They were water dwellers. Amphibians released and hatched their eggs in water. They spawned hundreds of tiny amphibians at a time. In reality primitive amphibians such Eryops (Cope, 1877) were food factories that produced food for themselves and for reptilians. Reptilians, which reproduced in small numbers, were mostly carnivores. They depended on the amphibia inside swamps for their sustenance. For self preservation and because of a need to protect their young many amphibians were large and aggressive. You can bet your bottom dollar that in the wee hours of Paleozoic and Mesozoic time many a large reptilian succumbed to the powerful jaws of large amphibians. Because small amphibia were a strong lure for reptilians, large amphibians in turn took advantage of this proximity and the hunter often became the hunted. But the bottom line here is that without the amphibians early evolution could not have progressed. The wonderful amphibians were entrusted with the task of converting zooplankton and phytoplankton into protoplasmic mass for the carnivores that had begun to occupy the land. This they did through hatchlings that were passive feeders and that later hunted tiny insects. Dimetrodon would have taken one heck of an empty bite at the plankton-filled waters of an early Permian swamp had it not been for the great amphibians. For centuries the importance of primitive amphibians has been overlooked. Amphibians are referred to as primitive creatures that went extinct. This extinction came only after sequential uplifting created watersheds, streams and rivers. Most swamps became buried. Latitudes changed. Both prey and predator began to occupy the land. Many types of fishes replaced the amphibia that once sustained the carnivores. Kudos to the great amphibians!
Ichniotherium cottae Footprints
Above: Ichniotherium cottae's footprints recorded in thick clayey mud. Notice that claw impressions merged with digit impressions creating the illusion of thick blunt digits. Soft thick mud is one of the major factors that impedes the accurate identification of footprints of the dim past. These "clawless" footprints are the muddy version of the footprints in the A to D group below.
Above: Ichniotherium cottae random manus-claw prints suggesting standing up and then getting back on all fours. Sounds unusual but there is ichnological evidence that Ichniotherium cottae lifted its front body atop low-lying branches while grazing on seed and spore appendages. This tetrapod possessed side-curved claws in the manner of the sloth and may have used them to hold on to vegetation. Before this elucidation in the El Pueblo early Permian swamp facies (Ichnonere, turning footprints, H. Ortiz 2014) the claw marks were previously believed to be swimming traces. Each background square is one inch scale.
Ichniotherium cottae Footprints as Key
Environmental Fossils of the El Pueblo ichnotelmafacies

______________________________________________________
Palaeosauropus in the El Pueblo
Early Permian Swamp Site
 |
Palaeosauropus ElPueblus (local
name only) Henry Ortiz 2013 |
Forebear of Chirotherium? Chirotherium (paper, D. Moor, '73), the human-hand beast of the Triassic, was deemed to be taller from the back and to have a thumb on the outer side of the pes or manus. The thumb probably increased traction on muddy surfaces. The human-hand-like footprint above (unknown if pes or manus) is Paleosauropus (Lea 1849). It had previously been found in the Carboniferous but apparently not in the Permian. Here's a Palaeosauropus footprint in the early Permian swamp-trajectory (the El Pueblo swamp facies) of northeastern New Mexico. Survival of tetrapods from the Permian to the Triassic is often debated because of rampant end-Permian volcanism. Did an early Permian precursor of the "hand beast" survive to become Chirotherium or did the early Triassic re-engender many species?
The Difficulty of Identifying Early
Permian Footprints
Paleontologists of the past such as Genitz, Pabst, Lea, Muller, and Marsh, to name a few, made incredible strides in naming different tetrapods of the early Permian. Nonetheless many ichnospecies and individual taxa have been misnamed in recent times. I myself keep confusing Dromopus with Tambachichnium (Muller, 1954). My confusion arises from the fact that infantile tetrapods were very lightweight and often imprinted only two digits, usually those on which more weight was exerted. One of my quandaries is, why am I unable to find the definitive adult counterpart of Dromopus (Genitz, 1861) or the infantile version of Tambachichnium? One of the most stable and reliable methods for positive identification, including the basic differentiation of reptilia from amphibia, is the correlation of adult and infantile footprints. While the above were probably two different types of tetrapods the criteria for better identification has not been met. The three main prerequisites for paleoichnological identification are adult and infantile footprint combinations; pes and manus combinations, and adult-to-infantile footprint ratio. The latter determines whether the taxa was reptilia (lower count) or amphibia (higher count). Already four and five digit manus footprints have been attributed to Tambachichnium. Which is it? And if indeed Dromopus (which simply means running-like tracks) is a smaller amphibian than Tambachichnium what is the adult size of its pes and manus? Somewhere I saw a revision of a Dromopus footprint that looks quite alike that of Tambachichnium. At least I have company for my confusion. I also saw a revision of Ichniotherium cottae (Pabst 1895) with a change from reptilian to amphibian. The revision is educated and I have often wondered if Ichniotherium cottae, because of its high count of adult footprints inside early Permian swamps, was not an amphibian. But the relatively small numbers of infantile footprints point to a reptilian. The investigation into the paleoichnology of the dim past continues with a luggage of many uncertainties. A lack of the proper fossils is usually the stumbling block for better identifications. Great progress has been made and future generations of paleoichnologists will hopefully refine the ichno-identities of all early Permian tetrapods.
Revisiting Standing-Lizard Footprints
(Henry Ortiz, 2000)
 |
| Another find (3/15/19) |
Above photograph: Footprints of Tambachichnium as further-defined ichnotype, Starelacerta Elpueblus. In 2000 I discovered a peculiar set of tetrapod footprints in the EPEPSF. The unusual quality of these footprints, that I would think are of the pes, was their symmetrical position, as when an animal or a human is standing still in observation of something. For a while I thought I had discovered the first bipedal proof from the early Permian. As I tried to sell this set of footprints a paleontologists quickly rebuked my finding, claiming that these were the footprints of a quadrupedal animal, and further added that these were the footprints of Tambachichnium, first discovered in the Tambach region of Germany. The paleontologist was possibly right on both counts. But by some miracle I had been modest in applying a name. Starelacerta merely means standing lizard, and not walking-on-two-feet-lizard. Stare in Latin means to stand; lacerta means lizard. My asking price for the Starelacerta fossil was ridiculously high, so of course, it didn't sell. Thank eternity. I recently found another set of the same type of footprints in the same standing position. These ones are a bit smaller. Three is better than one when it comes to evidence and here they are; Three similar sets of footprints poised in a standing position. Yes indeed, Starelacerta routinely stood on two legs, either to reach higher, or to gain better visual reference. And where's the trackway of a Tambachichnium, anyway? Some mystery here. Did Tambachichnium hop? One thing is certain. These footprints now comprise the first conclusive evidence of a bipedal stance being employed during the early (Artinskian) Permian Period. The bolosaurids were bipedal, though. So was Tambachichnium simply a bipedal bolosaurid? As an herbivore did it condition itself to stand erect in order to reach higher as it fed on ferns and other vegetation? More study needed here.
Edaphosaurus's Footprints
Edaphosaurus (Cope, 1882) was much larger than Dimetrodon and has often been described as the herbivore counterpart to the carnivore Dimetrodon. Edaphosaurus was also bigger than Ichniotherium cottae. Early Permian swamps and intermittent flowways between outlier swamps and along swamp trajectories leading from the upland, were well vegetated environments that necessitated direct herbivore control of many superfluous plants. Flowways could become excessively marshy and divert. That's where such giants as Edaphosaurus and Ichniotherium cottae came in. Both Edaphosaurus and Ichniotherium weighed in at several hundred pounds, although the EPEPSF species were of an earlier and smaller size. While no one has dared to identify the footprint of Edaphosaurus with any amount of certainty the footprint below is offered as a highly probable pes of the giant plant eater of the early Permian. New ichnological evidence showing a larger claw spread than that of Ichniotherium cottae has been quarried in the El Pueblo early Permian swamp facies. By natural ichnospecific count as outlined by the master paleontologist Edward Drinker Cope from skeletal evidence, the early Permian included Edaphosaurus as one of its most important herbivores of the time. This identification is based on the offset digit span that increased balance for pelycosaurs, and on the curved claws, which trampled light vegetation and conversely, served for the hooking and lowering of fronds and branches for grazing. (Update, 8/14/18) Recent finds of pes footprints attributed to Edaphosaurus in the EPEPSF exhibit four digits on pes and five digits on manus. This difference in digit count, plus elongated plantar regions on Edaphosaurus footprints may be used as the criteria for distinguishing between the footprints of Edaphosaurus and Ichniotherium cottae. Side-ward curved claws are standard characteristics for both species, and for other large herbivores (Henry Ortiz, 2018).
 |
| Pes footprint belonging to Edaphosaurus, an herbivore. |
The above footprint has been revised to a pes footprint because comparison to Ichniotherium cottae's footprints reveals that the plantar (main swivel pad) was oblong in Edaphosaurus and more rounded in Ichniotherium cottae. In general both ichnotypes point to short feet with long side-curved claws. Why not a long pes plantar region in both herbivores like in Dimetrodon, the pelycosaur carnivore? Probably because the large long claws of these herbivores necessitated a short roundish plantar region with a certain amount of back and forth roll that could lower or elevate the claw sets in relation to a hard or soft-mud surface.

Above photo shows a pes-manus set of Edaphosaurus's footprints. The much larger pes footprint suggests that this track maker was heavier toward the back, possibly for more agility of the front limbs. The pes claw-spread in this specimen is eight inches wide. Inwardly curved claws were designed so as not to hook on vegetation while walking. The curved claws create the illusion of a crescent step. The step of large herbivores was actually quite direct and designed to push thin vegetative stems out of the way (Henry Ortiz, 2017).
 |
Edaphosaurus manus footprint, 5-1/2 inches wide.
Claws did not record. |
Update: Due to the discovery of more probable Edaphosaurus footprints, it now appears that the smallish manus (above), which looks out of place for a large herbivore like Edaphosaurus, can now be explained. There is evidence that Edaphosaurus trackway was considerably wider than Dimetrodon's. Because of its large temperature regulating sail Edaphosaurus was wind-compromised, just like Dimetrodon. But Edaphosaurus dealt with the issue of side balance in a different way. The pes claw-spread was very wide but the manus claw-spread was not as wide. However, it appears that the manus limb-trackway width was wider than at the pes. And because Edaphosaurus El Pueblus's head was quite small, frontal weight was not that great. In both Ichniotherium cottae and in Edaphosaurus's footprints the plantar regions are small but one very important factor cannot be ignored: the curved claws were part of the weight-bearing design. That explains the superfluous amount of claw imprints in the EPEPSF. Although the Edaphosaurus of the EPEPSF was larger than IchnoDimetrodon El Pueblus an important ichnological fact continues to present itself: IchnoEdaphosaurus El Pueblus was larger than IchnoDimetrodon El Pueblus but was not a six-hundred pounder. Ichno-Dimetrodon of the EPEPSF weighed in at about seventy-five pounds. IchnoEdaphosaurus El Pueblus and Ichniotherium cottae el Pueblus weighed in at about (revision 27/18) one-hundred-fifty pounds, maximum. These sizes and weights fall in line with the EPEPSF Artinskian early Permian designation of 285 million years ago. It was many millions of years later that the six hundred pound Dimetrodon grandis and the giant Edaphosaurus came into existence because of a constant and adequate food supply close to the equator. Geological age estimates from surrounding ichnofacies were erroneously misinterpreted by previous researchers as being of Guadalupian age because of an insufficient number of fossils. At the Pennsylvanian/Permian boundary related to the EPEPSF, footprints of unknown tetrapods, mostly claw marks, are even smaller in general. Based on footprints, the Edaphosaurus of the EPEPSF was bigger than the Dimetrodons from same site. The 600 pound Edaphosaurus cruciger (Cope, 1878) found in Texas and Oklahoma is probably a younger version (further developed through evolutionary time) than ichnoEdaphosaurus elPueblus. In evolutionary time, younger is usually bigger.
Origins of Sauropods in the El Pueblo
Early Permian Swamp?
(A)
(B)
(C)

Early in 2018 the EPEPSF produced what may be herbivore footprints different from those of Ichniotherium cottae and Edaphosaurus. The plantar areas are roundish, symmetrical, and obviously, mud-sink resistant. Footprint (A) is a convex pes impression and is seven inches across digit spread. Both pes and manus appear to be four-digit. Footprints (B) and (C) are manus footprints and measure six inches across digit spread. It is possible that these footprints represent the early Permian origins of Sauropods. Even if the neck of an Antesauropod (Ortiz 2018) could reach only four feet high, the trend for trimming tall vegetation was possibly being set since the early Permian. The deviation from short to long-necked tetrapods would have entailed a constant evo-conditioning (focus of diligence) of attempting to reach for higher and higher vegetation. Both Ichniotherium cottae and Edaphosaurus qualify for this type of branching because of their endless pursuit of seed fruits that were usually located at higher spots in most vegetation. In confirmed sauropods of the Mesozoic, pes footprints appear roundish and with very short and oddly formed digits. These roundish (or wedge-like, front) footprints are the result of the massive weight that blended the entire physique of the pes and manus into a generalized depression. The manus footprints of giant sauropods are shorter than the pes, suggesting that the ichnological roots of the first definitive, and smaller, early Permian sauropods (with extra neck-length) will fall along those lines. Yet the manus footprints are or will be found to be roundish in Antesauropods because of extra neck length and longer than usual forward counterbalance.
Biggest Footprint Yet in the EPEPSF

Above photo: Revision note: This footprint, which measures eight-and-a-half inches across the digit impressions, remains indeterminate pending discovery of similar-size footprints. The symmetrical stubby digit or claw ends have cast doubt on an herbivore entity. This pes footprint somewhat resembles Ichniotherium sphaerodactylum (sphere-like digit ends, Pabst, 1895), an herbivore footprint common in the EPEPSF, but of a much smaller size, and with a front-to-back-compressed appearance. Unlike the above footprint, the latter mentioned footprints exhibit stubby digits with side-curved claws, and will be posted later. The other and next logical ichnotype possibilities based on an unusually large size are Sphenacodon or Ophiacodon, reptilians of the early Permian that grew to over nine feet in size (Marsh 1878). I would imagine that during Artinskian time the above-mentioned species would have been smaller, unless the above entities were the giants of the Artinskian Age. Hopefully more footprints of this size will be found in the EPEPSF in the interest of a better ichnotype confirmation. This footprint seems to point to an early Permian slope and in-swamp hunter (omnivore?) of obvious success, based on very large size.
Partial Footprint of Ichniotherium sphaerodactylum
Above photo: This pes footprint of Ichniotherium sphaerodactylum is lacking two of its longest digits. Ichniotherium sphaerodactylum was an herbivore and a smaller version of its early Permian contemporary, Ichniotherium cottae. Ichniotherium sphaerodactylum footprints are noted for rings around the digits, as visible above on two of the digits. The three main herbivores, including the two mentioned above plus Edaphosaurus, point to plenty of vegetation during the Artinskian early Permian. (It must be noted that some paleo environments were more lush than others, and in some impoverished environments a smaller taxanomic count may have been the result.)
Other Traces from the El Pueblo early
Permian Swamp Facies
 |
| Millipede traces |
Above photo: This is a key fossil of centipede traces. Key fossils clarify faint or subtle traces with clear traces. The clarification here is how millipede or centipede traces appear in watery mud, when individual pod marks are not recorded.
 |
| Plant profile about 4 inches wide. |
 |
| A rare find: traces of amphibian eggs |
Above shapes have been revised from indeterminate to amphibian egg impressions (Henry Ortiz, 2017) because they were found in a hatchery zone. The hatchery zone contains hundreds of newly hatched Dromopus sp. amphibian footprints. It is probable that these eggs were buried by successive sediment before they could hatch. Scientists have wondered since a long time what early Permian eggs looked like. They correctly deduced that eggs of the Permian were soft and that they were probably deposited on moist or wet surfaces because of the intense heat of the Period. Part of that question has now been answered. Their gelatinous-like shape was similar to frog eggs.
Above: Supporting fossil of amphibian eggs. The tiny thin impressions trailing from concave impressions are the traces of the formative-limb ends of newly hatched Araeocelids, probably Dromopus isp. The amphibious thin lizards (judging from their thin footprints) comprised the Dromopus
family and other amphibious groups. Fetal stage of Dromopus sp. was apparently salamander-like in appearance. The
immediate ichno-locality of these egg traces is rife with various age stages of
the Dromopus family. (Please note that ichnoDromopus has long been classified as a reptilian. My hypothetical classification of ichnoDromopus as an amphibian is based on the salamadroid entity and the evidence of schools of dromopus in small opportune shallow ponds, like tadpoles. Dependence on water because of the hot climate possibly promoted a tendency for underwater breathability in a majority of swamp-age animals [Henry Ortiz 2010].)
A Paleonychiological Find


Above photos: The sideways claws of this Dromopus marsh (Marsh 1894) manus or pes were physiologically similar to those of Ichniotherium
cottae. The similarity is based on the fact that both walked on sideward-positioned
curved claws. Why was that? First, tetrapods of the early Permian were unable to retract their
claws. Secondly, walking on a hard surface with erect claws of high curvature placed
a lot of pressure on the phalanges. So why not walk with claws in a sideways
position, and then when the need arose, such as for added traction in mud,
arenite, or when rummaging through leaves, turn them into a vertical position. This was probably accomplished through wrist-like action. This natural design of claws evolved in response to the different walking
surfaces of early Permian environments. Tetrapods walked on a variety of surfaces including very hard
dry ground, underwater concretions, or in deep soft mud, humus cover, or fine sand. This is the first-ever ichnological glimpse of Dromopus claws and distal phalanges. This find constitutes an important addition to Paleonychiology, the study of ancient
fingernails as claws, a concept introduced by Henry Ortiz in 2014. Left side segment is one inch and a half long. Note very sharp claw ends.
Skin Impression
Ichnology occasionally renders outer texture impressions such as plant bark, tree bark, or animal skin. The skin texture shown in above photo is from two of Tambachichnium's long, curved digits. The skin was similar to that of a modern lizard, chicken or bird; simply a pliable universal outer covering that was effective since Periods past to the present.
Identifying the Dromopus group is not an easy matter. The main obstacle is that Dromopus sp. footprints are almost always partially imprinted. Apparently, Dromopus ichnotypes were thin and lightweight. They exhibit a peculiar way of walking which exerted more weight on the front of their sideways claws. Note photo (1) where the claws have been poised in a more erect position. In photo (2) the claws are completely sideways. The identifications are as follow, although there is that possibility of error through partial recording in these identifications too: 1, 2 and 4, Dromopus agilis; 3, Dromopus lacertoides (Geinitz, 1861); 5, Dromopus marsh (Marsh 1894). Footprints vary from one-half inch for the smaller ones to one-and-a-half inch for the larger ones.
_______________________________________________
Might this be the adult Dromopus marsh footprint? Image shown above is seventeen inches long, demonstrating that an adult Dromopus marsh was a sizable tetrapod. Same-side steps are eight inches apart. Most footprints documented world-wide are infantile or of half-grown track makers traveling or milling around in groups. Why Dromopus marsh usually imprinted one side of its trackway much more than the other is a mystery yet unanswered. It is possible that Dromopus applied more weight to one side of its body as it carried its head to one side to assure front and back visual reference. The Dromopus group was the most common prey for reptilians but was comprised of the most active and picturesque herd animals of the early Permian.
Top photo: These are the footprints, possibly adult size, of Dromopus agilis. Dromopus agilis's mode of ambulation is still not fully understood because of the many imprints on a small area. This very agile tetrapod earned the suffix "agilis".

Above photo: Enlargement of Dromopus agilis pes and manus. Manus was considerably smaller than the pedes. This suggests much use of pedes, perhaps to the point of Dromopus agilis intermittently standing on its hind feet. The large pedes also suggest that Dromopus agilis was heavier from the back and might have sported a hefty tail. Although the further evolution of Dromopus agilis remains unknown, it was the repetitive use of the pedes in attaining an erect stance that enabled some tetrapods to become bipedal. The forelimbs became tiny and the tail large, providing the proper balance for bipedal mobility. Early Permian sites such as the El Pueblo facies were the evolution trial grounds of different bipedal dinosaurs of the future and one likely candidate of early bipedalism was Tambachichnium starelacerta, discussed in this blog (H. Ortiz, 2019).

Above photo: It's official: Dromopus marsh footprints and raindrop impressions are the most common trace fossils in the El Pueblo early Permian swampfacies. Please note that so far no concurrence has been found between depositional environment and ichnotypes within the intermediate slope of the El Pueblo trajectory. The only differences so far in the El Pueblo swamp facies are interchangeable and based on water depth, amount of vegetation, or supply of small amphibious prey. A noticeable difference does occur within the bottom-land alluvial-clay valley where a presence of medium and small amphibians was dominant. This stands to reason because of a watery environment. Already over seventy different types of footprints (representing over seventy different ichnotypes within a three acre area) have been found in the El Pueblo early Permian swamp facies. (During its functional existence the El Pueblo early Permian swamp was much bigger, with expansive gradual shores and surrounding vegetation.) This shows that swamp life of the Artinskian early Permian was varied and numerous. This tells us that first and foremost the El Pueblo early Permian swamp facies was a gathering point, a "metropolis" as some researchers refer to points of high tetrapod-type congregation. This gathering point was a multi-function environment that included a feeding source, spawning areas, and an adequate supply of water and vegetation. It is a well known fact that tetrapod types of the early Permian number in the hundreds and many researchers forego any attempt at a comprehensive cataloging of same, citing environmental-age variations to which some ichnotypes might be unique or exclusive. Part of the so called Permian extinction was the result of changing environments, something that becomes evident in the differing footprints from the early to the late Permian. Exhibiting clear co-relationships between prey and predator and between reptilia and amphibia, the EPEPSF represents one of the best core counts of classic early Permian footprints in the world. The life span of the El Pueblo early Permian was no more than forty years based on rainfall repetition and sedimentation rate, a very short geological time which confined floric and faunal trace evidence within a very tight coeval time frame. A short time frame for the El Pueblo facies is further reinforced by the fact that fossiliferous strata is confined to a depth of only four feet. This "screen shot" of geological time occurred through the incidence of uplift and spot subsidence, the modus operandi of the Permian Period that constructed multi-level terrestrial environments. Of natural necessity for the furtherance of evolution, terrestrial environments across equatorial Pangea contained water catchments such as the EPEPSF (Ortiz, 2018).
Was Evolution Rate Variable?

Above photo shows a small pes footprint (above number two on ruler) of the amphibian Cincosaurus from the EPEPSF. Its discovery in the early Pennsylvanian Period (Aldrich, 1930) and now in the Artinskian-early Permian Period reinforces something that I have been contemplating for some time: the late Carboniferous and the early Permian were fast-paced Periods because of accelerated rates of uplift and sedimentary disposition. The relatively fast speed of transformation from inland marine to terrestrial swamps was inevitable and is exemplified in the mechanics of tectonic collision: Transformation or modification occurs much faster during a plate collision than before or after the collision. Thus the late Carboniferous-to-early Permian transition occurred faster relative to less eventful and more drawn-out Periods of non-collision. Although continents pushed against each other very slowly, uplift and alluvial aggradations, the first driven by volcanic forces; the other by wet-tropical rains, were both relatively fast-paced. That same geological frenzy might explain the extensive grouping of tetrapods that congregated, evolved, devolved or went extinct in a relatively short time (Ortiz, 2019).
Interesting and Colorful Fossils
from the El Pueblo Quarry

Above: Enlarged photo of possible stem buds or spore cases. Note flower-like shapes.
Above: Rare tetrapod footprints on a post-rainfall wave. Note raindrop impressions from waning rain.

Above: These are multi-directional waves from ingress of flood water into the El Pueblo early Permian swamp. Dual-colored layers such as this are from the strata above the fossiliferous redbeds. This layer marks the beginning of final burial of the El Pueblo early Permian swamp. As the swamp began to disappear beneath colossal amounts of clayey alluvium, tetrapods, many in a more advanced stage, migrated further down the swamp valley ahead of an ever increasing onslaught of sediments from the upland. Swamps continued to be short term habitats which were buried and relocated at the whim of continual uplifting, monsoons, and dislodgement of sediments from the upland. The alluvial clay valley persevered as the vital location for survival throughout the rest of the Permian Period, with neo-Triassic dinosaurs evident only twenty-two miles down river. This shows that early Permian-to-Triassic evolution trajectories were much more compact than previously thought.
Flora From the Time of Dimetrodon
Above photo: A rare trace (colorized) of Autunia fern (Krasser 1921) in the El Pueblo facies. Autunia ferns were probably resistant plants and not the favorite plants for herbivores of the early Permian. Fronds are around three inches long, probably of a smaller plant. Nonetheless, the flora of the El Pueblo swamp does suggest a high level of resinous content in part from high levels of carbon dioxide. The many eatable-fern traces of the El Pueblo swamp exhibit a concentration of carbon toward the bottom of the stem, making the roots highly resistant to ground decay (Henry Ortiz, 8-2020).

Above photo shows a small amphibious trackway and a stem of Chaloneria (Pigg and Rothwell 1983). Chaloneria was a branchless long-stem plant with small open-scale leaves. Here the trace represents a flood-flattened Chaloneria embedded in mud. Flood-flattened muddy plants are the norm of vegetative traces in the EPEPSF. That's because the swamp depended on influxes of muddy water for its existence. In light of the high decay factor of the El Pueblo swamp now we know that Chaloneria was carbonically resistant to decay, and like most Permian plants and trees, probably stored water. (Henry Ortiz, 2020.)

Above colorized photo shows a scale-tree shoot of unknown type of early Permian tree. The interesting feature here is of coarse the bulbous top, a very common paleobotanical feature of the early Permian. Plants and trees strived to store water because of variable terraqueousy. The scale structure allowed for growth and outward "flowering". The top scales developed into large scales or fronds just like the shoots of modern coniferous trees.
Above photo: Colorized red-bed fossil showing small Walchia conifer cones from 285 million years ago. Fossil is five inches across at bottom. I'm inclined to associate smoke with fire and the local pine trees of the EPEPSF with the Walchia piniformis (Sternberg, 1825) shown above.

Above photo was colorized from EPEPSF leaf-mat fossil. It shows scaled leaves of Lebachia
hypnoides (Brongniart, 1801-1876). Instead of the small sticker spines of the Walchia piniformis, the individual leaves have a braided appearance. In my opinion these leaves are the ancestors of the local junipers, shown in the photo below.
The El Pueblo early Permian swamp facies "El Ichnofacies Clásico" is the long juniper-covered hill at top right. In the background is part of the Sangre de Cristo uplift. The arroyo in middle of photo leads to the namesake village of El Pueblo, once home to the Morriquez Puebloan Indians.



Above three photos show a long sinuous swamp plant from the El Pueblo swamp facies. The aquafloric transition from the Pennsylvanian to the Artinskian early Permian shows a high presence of clusters of awl or scale-like needles in aquatic plants. Descendants of the first shore mosses of previous Periods, these choice of needle leaves was meant to minimize evaporation during low-water exposure to the sun. Early Permian swamps, born on the ever-rising slopes of an uplifting landscape, often experienced sedimentation influx or rapid drainage that changed swamp-bottom elevations in a hurry. Narrow image (click to enlarge) shows form of one needle-like leaf cluster at corner of ruler.
The Layers of Time
The footprint fossil and the old separating wedge shown above were found together atop a pile of tailings. "Old time" and "new time" bring to mind the theory of circular time from whence we merely borrow "moments in time".
Elemental Dynamic Fossils
Besides trace fossils that were made by downward pressure from animal feet, trees or plants many fossils were made by elemental forces. Some of the most common are roll fossils left by rolling objects, such as dry plants. The markings below ruler in above photo are from scale leaves, possibly from a cycad cone or trunk being rolled along by a shallow current, wind, or both. The lines above and below the cycad marks are from plant roots or leaves dragging on the soft clayey mud as shallow current conveyed organic detritus into swamp. But the detritus was not waste or debris by a long shot. It provided valuable mulch and humus for the enrichment of the swamp substrate.
Beginning of the Triassic Period
Note: The following was slightly revised due to the discovery of more sill-volcanic evidence suggesting a higher production of calcium carbonates than previously thought.

View from south of the El Pueblo early Permian swamp facies. The EPEPSF is located to the left of tallest mesa at about 250 feet above Pecos River floodplain. In the distance is part of the Sangre de Cristo uplift, twenty-five miles away. Tallest mesa is about early Triassic. The two thick white stratifications on right-side of photo are quartzite lake-bottom formations enveloped and interspersed with calcium carbonate from very hot evaporation. Another clue to the initial volcanic-venting calamity that occurred around the end of the Permian, and that sometimes delivered heated water and steam through the bottom of thick stratifications. At some point in time, perhaps at the Permian-Triassic boundary, steam may have been rising from these white formations which were already concreted, and at level with the white stratifications on the opposite side of the alluvial clay valley. Note that alluvial-clay valley refers to a higher ratio of clay deposition and or smaller swamps of lesser concretion that more readily gave way to erosion.
__________________________________________________
Eventually the El Pueblo early Permian swamp became buried through massive sedimentation from the upland. The alluvial supply continued to be of the redbed type, indicating that there was no shortage of pink granite that was continually uplifting and weathering on the upland. Granite is an igneous product, a partial melt, if you will, of some of the most common elements on earth, including quartz, feldspar and trace amounts of iron and mica. Granite production is an ongoing process in an uplifting environment because of the enormous igneous activity that is precipitated by subductive plate movements that bring granitic material to the surface. Although a very durable material, granite is brittle. Granite chunks disintegrate quite readily as they strike other hard rocks during conveyance by strong water currents. Granite wash, which is a mixture of granitic sand, pebbles and other quartzitic material disintegrates or weathers into a fine arenite, which when mixed with organic particles makes up the soil around us. It was the finer sandy materials that slowly filled the inherent fault basins of the Upper Pecos Valley, creating the mesaland that is now visible over a thousand feet above the El Pueblo early Permian swamp. Larger chunks of granite and other rounded pebbles and cobblestones were deposited closer to the upland, a gravitational sediment-discriminatory process that never fails to amaze me. Above these mesas there are other mesas that add another three hundred feet or more of height above the El Pueblo early Permian level. The final level above both tiers of mesas consists of quartzite, which is a white to yellowish grainy but glassy material. The source of this quartzite material was just that; large chunks of quartz that broke free from huge granite blocks that became shattered through time, providing quartz sand for streams to convey over the mesaland. It is not far fetched to say that the supply of granite had ran out as uplifting waned during the late Permian. Weathering and runoff from the upland continued unabated. Fine quartzitic sands conveyed by runoff ended up inside mesa-level lakes. At the end of the Permian Period the mesaland, like other uniplanes before it, extended southward from the upland mountains now known as the Sangre de Cristo uplift.
 |
This ten-inch-across mushroom is fossilized in calcium
carbonate, no less. Talk about aridity! |
But a great mystery remains: what was the exact climate of the Permian-Triassic boundary. Many years ago a large ten-inch diameter mushroom fossil (shown above) was found below one of the mesas close to the El Pueblo early Permian swamp facies. Because mushrooms do not require too much sunlight this might imply that there was dimming of the atmosphere from volcanic activity. Above the mesaland a few miles to the south, a web-like pattern of igneous intrusion is present in rocks and boulders. This volcanic intrusion is from volcanic venting that intruded through cracks and crevasses into the thick stratifications that now form the tops of mesas. Thus, the eminent evidence is that a new and more severe round of uplifting and volcanic activity had begun, in essence, marking the Permian-Triassic boundary. Volcanic intrusion into mesa lakes, coupled with aerial dimming from smoke, might very well be the local root cause of the so-called great extinction of the end Permian. In an environment that was producing giant mushrooms, many different plants that necessitate photosynthesis may have been atrophied. Interestingly the growth of mushrooms would mean that the temperature was not blazing hot like in the early Permian, and in fact, had cooled down to about sixty to seventy degrees Fahrenheit. Small palm-tree forests (and or seed-fern trees?) were apparently thriving and also providing humus for mushroom growth via trees that fell inside mesa lake perimeters either through desiccation or high winds. Nonetheless, varied fossils of pro-ecological vegetation that was present during the Permian-Triassic boundary of the Upper Pecos Valley still remain to be found. As to the Permian-Triassic boundary climate, there were episodes of heavy rains and the sky was smoky from sill volcanic activity. There was a great spike in carbon dioxide throughout the boundary transition. The climate became more amiable as the early Triassic progressed, but the exact death toll from the local P-T boundary transition remains unknown.
 |
| Indeterminate or unnamed species |
Above: These two tree stems, the longest about nine inches long, are from Permian-Triassic-boundary (tentative) palm-tree forests. Their twirled and undeniable beautiful appearance suggest resistance and conditioning to very strong or steady winds. Or are these southwestern-like bark designs simply the patterns of circular growth? The forests were located inside broad lakes. The vestiges of these lakes seem to lack in vegetative variety. Mushrooms around the lakes may have been eatable. The only hope for more vegetative variety and the survival of animals of the Permian-Triassic boundary seems to point in the direction of the alluvial-clay valley, which is now vacated by massive erosion. The entire matrix is lacking in trace evidence from which to infer vegetative variety. Called bolillos (rolling pins) these palm-tree stem fossils were widely collected by old timers working above or below the end-Permian mesas of the upper Pecos Valley. And indeed, many the early settlers and the Natives used them on metates as rolling crushers or grinders.

These two tree stumps show growth rings recorded by a quartzite medium...

...yet some stumps are more prone to erosion than others. We might be looking at palm blade-leaf trees and or fern trees here, but without any traces of leaf patterns, these stumps remain speculative. Because different tree segments have been found embedded in individual thick slabs, it appears that this was a deep burial that occurred via several thick layers of quartzitic sediment. The slab-separating agent does not seem to be clay, but rather a buildup of calcium carbonates. A good guess here is that volcanic action itself, through the under-invasion of lakes, was producing a large amount of calcium carbonates. Large flora decreased, along with canopy protection. No wonder trees possessed a sparse canopy of leaves in order to minimize evaporation and sun blight. The fact that these were high bare-stalk trees in the manner of classic palm trees indicates water levels often rose, inundating mesa-level lakes. Sparse foliage also means high winds, although the cause of windy conditions at the local P-T boundary is uncertain. If we base a theory on the known evidence of high winds in the El Pueblo early Permian swamp that source was the paleo-west coast, from whence blew very strong tropical storm winds. If there was a presence of sustained winds that can only mean an appreciable increase in ancestral uplift, perhaps an apex juncture of intercontinental collisional pressure, which in itself, would have triggered the widespread sill volcanism. The sumptuous conveyance of quartzite sands indicates periods of heavy rainfall. Something had set in motion an unusual assortment of weather events, and perhaps as the Sages of Old said, look up to the heavens when looking for signs of war or havoc. The rain-forest type of climate was still in action. The final deposition that finalized the burial of these forest lakes was a barrage of large rounded river rock dislodged from the upland after a neo-Triassic uplift/monsoon flood episode. An obvious increase of upland and intermediate slope gradient had occurred. It appears that by then the lake floors were already concreted, and covered by a layer of redbed alluvium. A new supply of alluvial clay slowly created a new plain that engendered taller early Triassic mesas, followed much later by the beginning of the great erosive phase.

Above photo shows sill-volcano gaseous intrusion right through a thick layer of clay at the bottom part of the mesa from whence the mesa landscape photo was taken. This intrusion was so hot that it melted the clay into a burnt ceramic-like bubbly mass that my naturalist brother used to refer to as a "gorgujo", or the gurgling from a very hot source. Sills of the same type of material are present on the sides of some of the other local mesas. There are no volcanic cones nearby, suggesting volcanic sill venting that probably originated below the upland and traveled along the plutonic fault line. It was this same type of igneous activity that often intruded below lakes, making anyone wonder how early life endured this inferno in the first place. But let's look at the aftermath: Twenty miles down river as the crow flies a fair diversity of Middle Triassic? dinosaur fragmentary skeletons, as well as many large amphibians, prove that life and evolution continued. Some, if not all, of the dinosaur skeletons are of the Crurotarsal type (those evolved from crocodiles), which reinforces my belief that all dinosaurs evolved from Dimetrodon's in-swamp replacements, the crocs and the gators. It is quite possible that the ichnological connection of the Triassic Crurotarsal archosaurs downriver can be traced back to the El Pueblo early Permian swamp facies. But only if by chance, some of the footprints can be ascertained as belonging to a crocodilian. The number of digits in an early Permian crocodilian's pes or manus is still up in the air because a crurotarsal has three digits, and a modern crocodile has five. The tricky part here is that the first crocodilians might have possessed five pes digits and three manus digits (revised to four pes digits from recently found footprints). One pes digit may have devolved.This would be in line with the necessity for less pes digits so as to facilitate rapid bipedal mobility in muddy environments. The footprints belonging to a possible crocodilian-like tetrapod were recently discovered, as expected, in flowways not too far from the EPEPSF. Apparently that's where early Permian crocodilian-like tetrapods were spending their time.
Copyright © 2017 by Henry Ortiz. All rights reserved.
_______________________________________







.JPG)








.JPG)
.JPG)






























































































































